
Význam a limitace zrakových evokovaných potenciálů ve studiu patofyziologie migrény
Significance and Limitations of Visual Evoked Potentials in the Study of Pathophysiology of Migraine
Dysfunction of the central nervous system is considered to be the primary factor in the pathophysiology of migraines. Neurophysiological techniques enable non-invasive testing of pathophysiological hypotheses of migraines. In this respect, Visual evoked potentials (VEP) are historically and currently the leading modality, even though the traditional assessment of VEP did not prove useful. Abnormal processing of visual information in migraines is demonstrated by reduced or absent physiological decline in VEP amplitude (habituation deficit) during prolonged visual stimulation. This review article presents a selection of studies with VEP for migraine, including recent studies using neural excitability modulation (transcranial magnetic stimulation, hyperventilation and light deprivation). The findings from studies of habituation to VEP in migraines can support the theory of reduced preactivation thresholds of the cortex in the interictal period, based on the predominance of inhibitory mechanisms due to thalamo-cortical functional dissociation. However, migraine is characterized by cyclic changes in cortical activity – there is no VEP habituation deficit in preictal and ictal phases.
Key words:
visual evoked potentials – migraine – pathophysiology – habituation
Autoři:
M. Bednář 1; J. Kremláček 2,3; Z. Kubová 2; R. Taláb 4
Působiště autorů:
Rehabilitační klinika LF UK a FN Hradec Králové
1; Ústav patologické fyziologie LF UK v Hradci Králové
2; Neurologická klinika LF UK a FN Hradec Králové
3; LF UK v Hradci Králové
4
Vyšlo v časopise:
Cesk Slov Neurol N 2013; 76/109(2): 162-167
Kategorie:
Přehledný referát
Souhrn
Dysfunkce centrálního nervového systému je považována za primární faktor v patofyziologii migrény. Neurofyziologické techniky umožňují neinvazivně testovat patofyziologické hypotézy u migrény. Zrakové evokované potenciály (Visual Evoked Potentials, VEP) jsou historicky i v současné době v tomto směru vedoucí modalitou, i když klasické hodnocení VEP se ukázalo být nepřínosné. Abnormní zpracování zrakové informace u migrény dokládá snížení anebo chybění fyziologického poklesu amplitudy VEPs (deficit habituace) na protrahovanou zrakovou stimulaci. Shrnující článek přináší výběr studií s VEP u migrény včetně recentních studií využívajících modulační postupy neurální excitability (transkraniální magnetická stimulace, hyperventilace, světelná deprivace). Na základě poznatků o habituaci VEP u migrény lze podpořit teorii snížené preaktivační úrovně kortexu v interiktálním období, a to na podkladě převahy inhibičních mechanizmů v důsledku funkční thalamo-kortikální diskonekce. Migrénu však charakterizují cyklické změny kortikální aktivity – v preiktální a iktální fázi nebyl zjištěn deficit habituace VEP.
Klíčová slova:
zrakové evokované potenciály – migréna – patofyziologie – habituace
Úvod
Migréna je stále chápána jako onemocnění, které je spojeno s hyperexcitabilitou centrálního nervového systému [1]. Výsledky neurofyziologických studií u migrény s významným podílem zrakových evokovaných potenciálů (Visual Evoked Potentials, VEP) však v posledních téměř třech desetiletích přinesly zásadní změnu v chápání neuronálních procesů spojených s migrénou.
V souvislosti s elektrofyziologickými nálezy u migrény začal být skloňován termín habituace, který byl do té doby v neurofyziologii zcela cizí. Habituace je obecný biologický fenomén, jímž se označuje pokles odpovědi na opakovaný podnět. V širším kontextu se jedná o jednoduchou formu neasociativního učení. Jde o aktivní proces na úrovni centrálního nervového systému, který by měl splňovat další předpoklady, např. proces dishabituace (promptní vymizení habituace, pokud je stimulace přerušena jiným podnětem) [2]. V běžné klinické praxi v neurologii, kdy je diagnostika VEP obvykle zaměřena na detekci demyelinizační prechiazmatické léze zrakové dráhy, je vyšetřován a zprůměrněn určitý počet odpovědí (obvykle 100–250) a výsledkem je jediná křivka. Pokud je záznam VEP rozdělen do více bloků, je možné sledovat pokles odpovědi (amplitudy VEPs) v čase jako reakci na protrahovanou stimulaci. Tento pokles bývá autory prací terminologicky ne zcela přesně označován jako habituace. Na poklesu amplitudy odpovědi na senzorickou stimulaci se nepochybně podílí i adaptace na receptorové úrovni a únava.
Po úvodu do problematiky VEP, který je pro další porozumění textu nezbytný, předkládáme výběr studií s VEP u migrény se zřetelem na interpretaci jejich výsledků ve vztahu k její patofyziologii. Zabýváme se také limitacemi a příčinami diskrepancí výsledků těchto studií.
Zrakové evokované potenciály (Visual Evoked Potentials, VEP)
VEPs jsou bioelektrické koreláty neuronální aktivity zrakového kortexu vyvolané jako reakce na přesně definovaný podnět. Jedná se o potenciály v řádu mikrovoltů, snímané povrchovými elektrodami ze skalpu nad zrakovým kortexem. Zaznamenaná odezva obsahuje také spontánní EEG aktivitu (obvykle potlačenou metodou zprůměrnění – averaging).
Přítomnost vysokých prostorových frekvencí s dostatečným kontrastem při zvratu světlých a tmavých polí šachovnice na obrazovce monitoru (Pattern-Reversal VEP, PR-VEP) u vyšetřované osoby vede k predilekční aktivaci makulární části sítnice a primární zrakové kůry [3]. Nad primárním zrakovým kortexem (oblast V1) je registrován výrazný trifázický potenciál, označovaný jako NPN komplex (negativní, pozitivní a negativní vlna). Jednotlivé vlny NPN komplexu se označují podle latencí (v ms), ve kterých se typicky vyskytují (N70 nebo N75, P100, N135 nebo N140, event. N145), někdy jen jako odpovědi N1_P1_N2.
Mezi základní parametry stimulace patří velikost čtverců šachovnicového podnětu, jas (luminance), kontrast a frekvence stimulace [4]. Velikost čtverců je udávána jako úhlová velikost v úhlových minutách (´) či stupních (1° = 60´ a odpovídá 1/360 kruhu) – jedná se o vyjádření úhlu, pod kterým vyšetřovaná osoba vidí stranu čtverce. Druhou možností, jak definovat velikost čtverců, je tzv. prostorová frekvence, která je definována jako počet čtverců, jež lze pozorovat pod prostorovým úhlem 1° (v anglické literatuře v jednotkách c/deg, resp. cpd – „cycles per degree“). Jas stimulačního pole je udáván v cd/m², kontrast v % jako podíl rozdílu a součtu jasů světlých a tmavých polí. Frekvence stimulace (v Hz) je dána počtem opakování stimulačního vzorce za 1 s.
Při užití stimulačního paradigmatu s pohybem struktury v zorném poli dochází k převažující aktivaci periferních částí sítnice (magnocelulárního systému) a navazujícího dorzálního proudu zrakové dráhy a extrastriátového zrakového kortexu (zejména area V5, mediotemporální oblast) [3]. Z praktického hlediska (relativně nižší interindividuální variabilita) se ukázala být nejvýhodnější reakce na začátek pohybu, tzv. Motion-onset VEP (M-VEP), jejíž hlavní komponentou je negativní vrchol N2 (s latencí kolem 160–200 ms). Vrchol P1 (s latencí cca 120–130 ms) naproti tomu odpovídá aktivitě parvocelulárního systému [3].
Charakteristiky VEPs jsou ovlivněny parametry použité stimulace. Charakter stimulačního podnětu je rozhodující pro cílovou aktivaci určité části sítnice a dalších subsystémů zrakové dráhy až po kortikální oblasti, což je nutné zohlednit v umístění elektrod (Oz pro PR-VEP, Oz/O1/O2/Pz u M-VEP, dle systému 10–20). Pro vyšetření parvocelulárního systému bývá doporučováno užití vysokého kontrastu a vysoké prostorové frekvence, pro vyšetření magnocelulárního systému (N2 odpověď při M-VEP) opačné parametry [5].
Při nižší luminanci a nižším kontrastu se snižuje amplituda PR-VEPs, stejně tak při nižší zrakové ostrosti [6]. V dospělosti se s věkem prodlužují latence a klesá amplituda VEPs. U M-VEP byl zjištěn významnější vliv stárnutí než u PR-VEP [7].
Omland et al ve své práci zjistili, že paradigma stimulačního podnětu u PR-VEP ovlivňuje habituaci VEPs. Habituace byla nejvýraznější při užití malých čtverců (8´) a relativně vyšší frekvence stimulace (3 Hz ve srovnání s 1,5 Hz). Při užití velkých čtverců (65´) nebyla habituace vyvolána vůbec [8].
Studie se zrakovými evokovanými potenciály (Pattern-Reversal VEP) u migrény
PR-VEPs se staly vedoucí metodikou ve výzkumu centrálního zpracování zrakové informace u migrény. Literární údaje o M-VEP u migrény prakticky chybí. Výjimkou je práce Szanyi et al, kdy byly u migreniků do studie zařazeny vedle PR-VEP i M-VEP, avšak design studie neumožňoval hodnocení habituace [9].
Detailní přehled studií s PR-VEPs u migrény podrobně uvádí přehledný článek J. Schoenena et al z roku 2003 [10]. Vyšetřování PR-VEP v klasickém schématu se ukázalo nepřínosné, výsledky byly značně rozporuplné (od vyšších po nižší amplitudy a/nebo prodloužení či zkrácení latencí). Byly používány velmi rozdílné metodiky, např. velikost čtverců kolísala od 8´do 50´, frekvence stimulace od 1 do 3,9 Hz, počet zprůměrněných odpovědí od 50 do 256. Soubory migreniků a zdravých dobrovolníků se pohybovaly většinou v počtu od 10 do 60, průměrný věk většinou mezi 30 a 40 lety. Hlavní limitací těchto studií se ukázalo být nedostatečné sledování faktorů, které mohly ovlivnit parametry VEPs. Jednalo se zejména o definici interiktálního období, resp. časového vztahu k předchozí a následné atace migrény.
Přelomovou prací se ukázala být práce autorů Schoenena et al z roku 1995, v níž byla poprvé užita metoda hodnocení amplitud PR-VEPs zprůměrněných v několika blocích, takže bylo možné sledovat pokles amplitudy VEPs ve vztahu k prvnímu bloku. Zatímco u zdravých dobrovolníků byla nalezena předpokládaná redukce amplitudy (a nazvána habituací), tak u migreniků byl nalezen deficit habituace, resp. dokonce vzestup amplitudy VEPs (potenciace) [11]. Následovaly další práce, které obecně deficit habituace potvrzovaly [12–19], i když ne všechny [20,21]. Důvody pro diskrepanci nálezů byly v rozdílné metodologii a v načasování vyšetření (preiktální vs interiktální fáze migrény).
V kontextu nálezů, které ukázaly, že během ataky migrény a těsně před migrénou nebyl zjištěn deficit habituace [15], začal být postupně kladem důraz na přesné definování interiktálního období (většinou 72 hod před atakou migrény a po ní).
Nižší amplituda iniciálního bloku VEPs u migreniků ve srovnání se zdravými jedinci vedla k interpretacím o nižší preaktivační úrovni zrakového kortexu, tedy o „hypoexcitabilitě“ v interiktálním období (v kontradikci s převládající představou hyperexcitability kortexu u migrény) [22]. Byla zjištěna korelace mezi amplitudou iniciální bloku VEPs a následnou velikostí habituace [18]. Nižší preaktivační úroveň kortexu pravděpodobně souvisí s deficitem habituace (zdraví jedinci podle této teorie snadno dosáhnou „stropu“ v aktivaci kortexu při opakované stimulaci a vykazují habituaci, zatímco migrenici „začínají“ na nižší úrovni aktivace kortexu, proto nedosáhnou tohoto „stropu“ a habituaci nevykazují) [22].
Deficit habituace VEP nebyl ovlivněn aplikací červeného světla (při použití barevných brýlí), které u jinak zdravých jedinců vede k potenciaci – což koresponduje s představou, že zrakový kortex migreniků v interiktálním období není zvýšeně excitabilní [23].
Bohotin et al užili repetitivní transkraniální magnetickou stimulaci (rTMS) zrakového kortexu, aby sledovali její modulační efekt na habituaci následně snímaných PR-VEPs. Vysokofrekvenční 10 Hz rTMS (facilitační) vedla k obnově habituace u migreniků, zatímco nízkofrekvenční 1Hz rTMS (inhibiční) neměla u migreniků signifikantní efekt. Tyto nálezy znovu podpořily teorii o bazální hypoaktivitě zrakováho kortexu migreniků v interiktálním období [24].
Světelná deprivace zvyšuje excitabilitu zrakového kortexu, a to cestou snížení aktivity („down regulace“) inhibičních GABAergních okruhů (GABA – kyselina gama-aminobutyrová). Jedná se vlastně o model deaferentace zrakového kortexu [25]. Palermo et al zkoumali vliv světelné deprivace a současně aplikované rTMS zrakového kortexu na habituaci PR-VEPs u zdravých jedinců. Zjistili, že vysokofrekvenční 10 Hz rTMS (excitační) dokáže restituovat defektní habituaci, která byla navozena světelnou deprivací. Tyto výsledky byly interpretovány v tom smyslu, že pro vznik habituace jsou podstatné právě inhibiční mechanizmy, významnou roli zde zřejmě hrají GABAergní okruhy [26].
Světelná deprivace u migreniků nevedla ke změně habituace PR-VEPs, došlo k významné redukci amplitudy prvního bloku PR-VEPs ve srovnání se zdravými dobrovolníky. Autoři uzavírají, že za deficitem habituace u migreniků nestojí inhibiční mechanizmy deaktivované světelnou deprivací [27].
Coppola et al zkoumali vliv hyperventilace na habituaci PR-VEPs. Hyperventilace u zdravých dobrovolníků vyvolala deficit habituace, u migreniků prohloubila preexistující deficit habituace. Autoři se v interpretaci odvolávají na předpokládaný inhibiční vliv neuronálních a chemických změn indukovaných hyperventilací (snížený mozkový průtok, zvýšená hladina laktátu, alkalóza, hypokapnie), a to primárně na subkortikální struktury (thalamus, retikulární formace mezencefala) [28]. Hyperventilace, která běžně provází stres, může být nahlížena jako provokující faktor vzniku migrenózního záchvatu. Paradoxně však právě bezprostředně před atakou migrény není zjišťován deficit habituace.
Analýza vysokofrekvečních oscilací VEPs v gama pásmu (Gamma Band Oscilations, GBOs) ukázala u migreniků redukci amplitud časných komponent (korespondujících s thalamo-kortikální aktivací) i sníženou habituaci pozdních odpovědí (reprezentujících postsynaptickou kortikální aktivitu). Autoři na základě těchto výsledků předkládají hypotézu, že funkční thalamo-kortikální diskonekce (hypofunkce serotoninergních drah) vede k nižší preaktivační úrovni kortexu, což indukuje deficit habituace [18].
Aplikace bolestivého podnětu u zdravých jedinců vedla k dishabituaci [29], zatímco u migreniků nikoli, což bylo interpretováno jako korelát hypoaktivity monoaminergních kmenových jader, která modulují thalamo-kortikální dráhy [19].
Obecně ve studiích s VEP u migrény nebyly zjištěny zásadní rozdíly ve výsledcích u migrény s aurou a migrény bez aury [10].
Diskuze
Příčiny určitých rozdílů ve výsledcích studií s VEP u migrény je nutné hledat v nesourodosti souboru subjektů ve studiích (migreniků i zdravých dobrovolníků, odlišností ve věku, v tíži a charakteru migrény, v délce trvání migrény, v rozdílném načasování vyšetření a kontroly interiktálního období, fáze menstruačního cyklu u žen apod.) a také v rozdílné metodice při vyšetřování VEP (parametry stimulace, způsob hodnocení, počet odpovědí v jednotlivých blocích apod.) – u vybraných studií detailně v tab. 1.
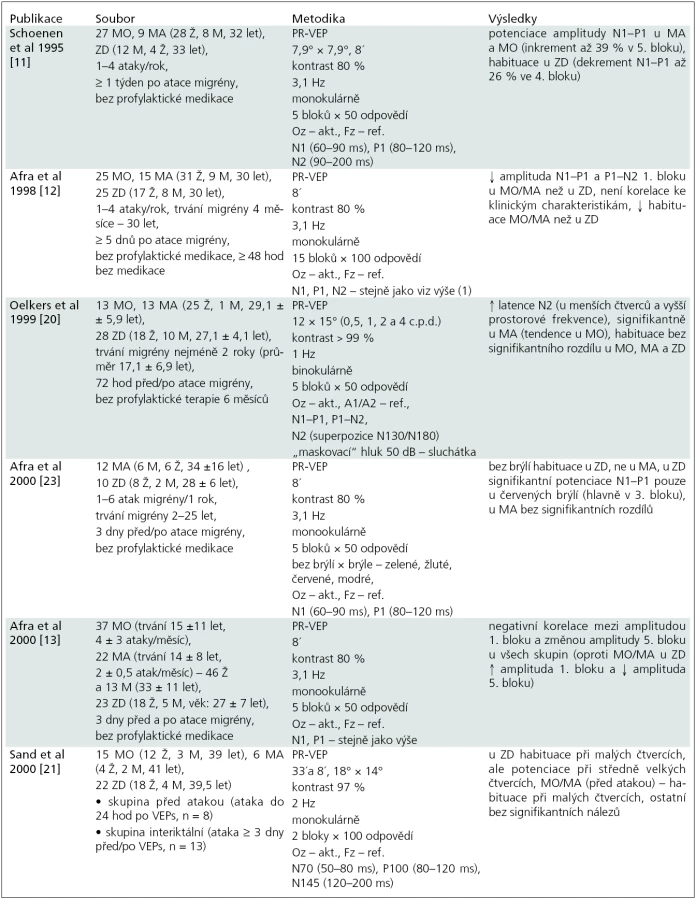

Hlavní limitací studií s VEP v aplikaci pro stanovení habituace je nepochybně množství faktorů, které na neuronální aktivitu kortexu v průběhu vyšetření působí. Některé z nich lze považovat za velmi obtížně ovlivnitelné – afektivní stav v době vyšetření, ospalost, ztrátu pozornosti [30], vliv nikotinu či kofeinu, vliv medikace, hormonální vlivy u žen apod.
Zajímavé je, že v žádné ze studií nebylo zohledněno užívání magnezia, které pacienti s migrénou často užívají (i jako volně prodejné preparáty). Přitom byl konstatován pokles amplitudy VEPs při zvýšení sérové hladiny magnezia, je-li u migreniků užíváno profylakticky [31]. V průběhu menstruačního cyklu anebo při hormonální substituční léčbě byly zjištěny vyšší amplitudy PR-VEPs při vyšších hladinách estrogenů [32,33]. Byl dokumentován také vliv medikace na habituaci PR-VEPs – profylaktická medikace betablokátory vedla k odstranění deficitu habituace PR-VEPs u migreniků [34], stejně tak medikace fluoxetinem ze skupiny SSRI (Selective Serotonin Reuptake Inhibitors) [16].
Nepříliš jasný je fyziologický význam fenoménu habituace v souvislosti s nálezy u migrény. Zdá se, že deficit habituace může působit selhání fyziologické ochrany mozku před senzorickým „přestimulováním“, což může hrát roli v patofyziologii migrény [35].
V žádné z prací nebylo použito přerušení zrakové stimulace jiným podnětem, aby byla skutečně odlišena habituace od senzorické adaptace a únavy, případně ztráty pozornosti či vigility.
Závěr
Neurofyziologické studie u migrény činí teorii prosté hyperexcitability centrálního nervového systému v současnosti neudržitelnou. Deficit habituace u migrény v interiktálním období je v současné době vysvětlován převahou inhibiční mechanizmů v důsledku funkční thalamo-kortikální dysrytmie (diskonekce). V preiktální a iktální fázi migrény není deficit habituace přítomen.
Deficit habituace byl u migrény opakovaně popsán napříč různými modalitami evokovaných odpovědí – u sluchových evokovaných potenciálů (Brainstem Auditory Evoked Potentials, BAEP), somatosenzitivních evokovaných potenciálů (Somatosensory Evoked Potentials, SSEP), u CNV (Contingent Negative Variation) a u kognitivních evokovaných potenciálů (vlny P300), ale také u blink reflexu – detailně v přehledném článku autorů Coppola et al [36].
Na druhou stranu fakt, že deficit habituace při různé senzorické stimulaci byl nalezen i u jiných onemocnění (např. u tinitu, schizofrenie, Parkinsonovy nemoci nebo u bolestí zad), vede k hypotéze, že fenomén deficientní habituace není specifický pro migrénu, ale je jen projevem chronicity onemocnění [37].
V budoucnosti lze očekávat propojení elektrofyziologických studií s výzkumem na úrovni molekulární genetiky a buněčného metabolizmu u migrény. Elektrofyziologické studie mohou pomoci v hledání a sledování účinku farmak v léčbě migrény, i když korelace mezi elektrofyziologickým efektem a účinkem léčby nemusí být vždy těsná (např. SSRI nepředstavují první volbu v profylaxi migrény). Vizí do budoucnosti je nepochybně i s pomocí elektrofyziologických studií ještě bližší poznání patofyziologie migrény s vyústěním do účinnějších terapeutických postupů (včetně využití efektu rTMS).
Autoři deklarují, že v souvislosti s předmětem studie nemají žádné komerční zájmy.
Redakční rada potvrzuje, že rukopis práce splnil ICMJE kritéria pro publikace zasílané do biomedicínských časopisů.
MUDr. Michal Bednář
Rehabilitační klinika
LF UK a FN Nezvalova 265
500 03 Hradec Králové
e-mail: michal.bednar@fnhk.cz
Přijato k recenzi: 14. 9. 2012
Přijato do tisku: 11. 12. 2012
Zdroje
1. Welch KM, D’Andrea G, Tepley N, Barkley G, Ramadan NM. The concept of migraine as a state of central neuronal hyperexcitability. Neurol Clin 1990; 8(4): 817–828.
2. Thompson RF. Habituation: a history. Neurobiol Learn Mem 2009; 92(2): 127–134.
3. Kuba M. Motion-onset visual evoked potentials and their diagnostic applications. Hradec Králové: Nucleus 2006.
4. Stejskal J et al. Evokované odpovědi a jejich klinické využití. Praha: Praga Publishing 1993.
5. Kubova Z, Kuba M, Spekreijse H, Blakemore C. Contrast dependence of motion-onset and pattern-reversal evoked potentials. Vision Res 1995; 35(2): 197–205.
6. Bartel PR, Vos A. Induced refractive errors and pattern electroretinograms and pattern visual evoked potentials: implications for clinical assessments. Eletroencephalogr Clin Neurophysiol 1994; 92(1): 78–81.
7. Langrova J, Kuba M, Kremlacek J, Kubova Z, Vit F. Motion-onset VEPs reflect long maturation and early aging of visual motion-processing system. Vision Res 2006; 46(4): 536–544.
8. Omland PM, Nilsen KB, Sand T. Habituation measured by pattern reversal visual evoked potentials depends more on check size than reversal rate. Clin Neurophysiol 2011; 122(9): 1846–1853.
9. Szanyi J, Kuba M, Kremláček J, Chlubnová J, Waberžinek G. Elektrofyziologické nálezy u migrény. Cesk Slov Neurol N 2001; 64/97(6): 349–354.
10. Schoenen J, Ambrosini A, Sándor PS, Maertens de Noordhout A. Evoked potentials and transcranial magnetic stimulation in migraine: published data and viewpoint on their pathophysiologic significance. Clin Neurophysiol 2003; 114(6): 955–972.
11. Schoenen J, Wang W, Albert A, Delwaide PJ. Potentiation instead of habituation characterizes visual evoked potentials in migraine patients between attacks. Eur J Neurol 1995; 2(2): 115–122.
12. Afra J, Cecchini AP, De Pasqua V, Albert A, Schoenen J. Visual evoked potentials during long periods of pattern-reversal stimulation in migraine. Brain 1998; 121(2): 233–241.
13. Afra J, Cecchini AP, Sándor PS, Schoenen J. Comparison of visual and auditory evoked cortical potentials in migraine patients between attacks. Clin Neurophysiol 2000; 111(6): 1124–1129.
14. Wang W, Wang GP, Ding XL, Wang YH. Personality and response to repeated visual stimulation in migraine and tension-type headaches. Cephalalgia 1999; 19(8): 718–724.
15. Judit A, Sándor PS, Schoenen J. Habituation of visual and intensity dependence of auditory evoked cortical potentials tends to normalize just before and during the migraine attack. Cephalalgia 2000; 20(8): 714–719.
16. Ozkul Y, Bozlar S. Effects of fluoxetine on habituation of pattern reversal visually evoked potentials in migraine prophylaxis. Headache 2002; 42(7): 582–587.
17. Di Clemente L, Coppola G, Magis D, Fumal A, De Pasqua V, Schoenen J. Nociceptive blink reflex and visual evoked potential habituations are correlated in migraine. Headache 2005; 45(10): 1388–1393.
18. Coppola G, Ambrosini A, Di Clemente L, Magis D, Fumal A, Gérard P et al. Interictal abnormalities of gamma band activity in visual evoked responses in migraine: an indication of thalamocortical dysrhythmia? Cephalalgia 2007; 27(12): 1360–1367.
19. Coppola G, Currà A, Serrao M, Di Lorenzo C, Gorini M, Porretta E et al. Lack of cold pressor test-induced effect on visual-evoked potentials in migraine. J Headache Pain 2010; 11(2): 115–121.
20. Oelkers R, Grosser K, Lang E, Geisslinger G, Kobal G, Brune K et al. Visual evoked potentials in migraine patients: alterations depend on pattern spatial frequency. Brain 1999; 122(6): 1147–1155.
21. Sand T, Vingen JV. Visual, long-latency auditory and brainstem auditory evoked potentials in migraine: relation to pattern size, stimulus intensity, sound and light discomfort thresholds and pre-attack state. Cephalalgia 2000; 20(9): 804–820.
22. Schoenen J. Deficient habituation of evoked cortical potentials in migraine: a link between brain biology, behavior and trigeminovascular activation? Biomed Pharmacother 1996; 50(2): 71–78.
23. Afra J, Ambrosini A, Genicot R, Albert A, Schoenen J. Influence of colors on habituation of visual evoked potentials in patients with migraine with aura and in healthy volunteers. Headache 2000; 40(1): 36–40.
24. Bohotin V, Fumal A, Vandenheede M, Gérard P, Bohotin C, Maertens de Noordhout A et al. Effects of repetitive transcranial magnetic stimulation on visual evoked potentials in migraine. Brain 2002; 125(4): 912–922.
25. Boroojerdi B, Battaglia F, Muellbacher W, Cohen LG. Mechanism underlying rapid experience-dependent plasticity in the human visual cortex. Proc Natl Acad Sci U S A 2001; 98(25): 14698–14701.
26. Palermo A, Giglia G, Vigneri S, Cosentino G, Fierro M, Brighina F. Does habituation depend on cortical inhibition? Results of an rTMS study in healthy subjects. Exp Brain Res 2011; 212(1): 101–107.
27. Coppola G, Crémers J, Gérard P, Pierelli F, Schoenen J. Effects of light deprivation on visual evoked potentials in migraine without aura. BMC Neurol 2011; 11: 91.
28. Coppola G, Currà A, Sava SL, Alibardi A, Parisi V, Pierelli F et al. Changes in visual-evoked potential habituation induced by hyperventilation in migraine. J Headache Pain 2010; 11(6): 497–503.
29. Coppola G, Serrao M, Currà A, Di Lorenzo C, Vatrika M, Parisi V et al. Tonic pain abolishes cortical habituation of visual evoked potentials in healthy subjects. J Pain 2010; 11(3): 291–296.
30. Kremlacek J, Kuba M, Kubova Z, Langrova J, Vit F, Szanyi J. Within-session reproducibility of motion-onset VEPs: effect of adaptation/habituation or fatique on N2 peak amplitude and latency. Doc Ophtalmol 2007; 115(2): 95–103.
31. Köseoglu E, Talaslioglu A, Gönül AS, Kula M. The effects of magnesium prophylaxis in migraine without aura. Magnes Res 2008; 21(2): 101–108.
32. Yilmaz H, Erkin EF, Mavioğlu H, Sungurtekin U. Changes in pattern reversal evoked potentials during menstrual cycle. Int Ophtalm 1998; 22(1): 27–30.
33. Yilmaz H, Erkin E, Mavioğlu H, Laçin S. Effects of oestrogen replacement therapy on pattern reversal visual evoked potentials. Eur J Neurol 2000; 7(2): 217–221.
34. Sándor PS, Afra J, Ambrosini A, Shoenen J. Prophylactic treatment of migraine with beta-blockers and riboflavin: differential effects on the intensity dependence of auditory evoked cortical potentials. Headache 2000; 40(1): 30–35.
35. Ambrosini A, Schoenen J. Electrophysiological response patterns of primary sensory cortices in migraine. J Headache Pain 2006; 7(6): 377–388.
36. Coppola G, Pierelli F, Schoenen J. Habituation and migraine. Neurobiol Learn Mem 2009; 92(2): 249–259.
37. Stankewitz A, May A. The phenomenon of changes in cortical excitability in migraine is not migraine-specific – a unifying thesis. Pain 2009; 145(1–2): 14–17.
Štítky
Dětská neurologie Neurochirurgie NeurologieČlánek vyšel v časopise
Česká a slovenská neurologie a neurochirurgie

2013 Číslo 2
Nejčtenější v tomto čísle
- Creutzfeldtova-Jakobova choroba
- Spinocerebelární ataxie typ 7 (SCA7) – kazuistika
- Lymeská borelióza jako příčina bilaterální neuroretinitidy s výraznou jednostrannou hvězdicovitou makulopatií u osmileté dívky
- Elektrofyziologické vyšetření pánevního dna