
Analýza „default mode“ sítě u zdravých dobrovolníků
“Default Mode” Network Analysis in Healthy Volunteers
The default mode network (DMN) is an organized network of brain structures involved in brain activity that may be observed in the resting state. In the course of the performance of an experimental cognitive task during functional MRI examination (fMR), these regions manifest as “deactivations”. The main areas involved in this network are the ventromedial prefrontal cortex/anterior cingulate cortex, posterior cingulate cortex/precuneus and angular gyrus/inferior parietal cortex. In a group of 10 healthy volunteers we employed the following approaches to the detection of DMN: deactivations related to a visual spatial memory task; seed functional connectivity from the specific region of interest (cluster posterior cingulate cortex/precuneus); and independent component analysis (ICA). We then sought correlations between the MRI signal and the results of the visuo-spatial memory task and the Addenbrook cognitive examination (ACE), in concrete terms with the ACE verbal fluency subscore (VFT), and memory. The ICA approach revealed a higher correlation rate with the results from functional connectivity compared with pure deactivation mapping. We found correlation between MRI signal in the cluster posterior cingulate cortex/precuneus and VFT performance.
Key words:
default mode network – functional magnetic resonance – cognitive task – deactivations – resting state
Autoři:
L. Krajčovičová; M. Mikl; R. Mareček; I. Rektorová
Působiště autorů:
I. neurologická klinika LF MU a FN u sv. Anny v Brně
Vyšlo v časopise:
Cesk Slov Neurol N 2010; 73/106(5): 517-522
Kategorie:
Původní práce
Souhrn
Default mode síť (DMN) je organizovaná síť mozkových oblastí zapojených do mozkové aktivity pozorovatelné v klidovém stavu. Při cíleném provádění experimentální kognitivní úlohy v průběhu vyšetření funkční magnetickou rezonancí (fMR) se tyto oblasti projevují jako tzv. deaktivace. Hlavními oblastmi zapojenými do této sítě jsou ventromediální prefrontální kortex/přední cingulum, zadní cingulum/precuneus a gyrus angularis/lobulus parietalis inferior. Pro sledování DMN u naší skupiny 10 zdravých dobrovolníků jsme použili jednak zobrazení deaktivace ve vztahu k paměťovému úkolu a korelační analýzu (tzv. seed funkční konektivitu) vycházející z oblasti zájmu přestavující cluster zadní cingulum/precuneus, jednak zobrazení pomocí analýzy nezávislých komponent (independent component analysis, ICA). Dále byly provedeny korelace fMR signálu s výkonem ve vizuálním prostorově paměťovém testu a v Addenbrookském kognitivním testu (ACE), konkrétně se subskóry verbální fluence (VFT) a paměť. Zobrazení DMN pomocí seed funkční konektivity významně lépe korelovalo s výsledkem ICA analýzy než s obrazem prosté deaktivace. Dále jsme našli korelaci mezi MR signálem v clusteru zadní cingulum/precuneus a kognitivním výkonem ve VFT.
Klíčová slova:
default mode síť – funkční magnetická rezonance – kognitivní úkol – deaktivace – resting state
Práce byla podpořena výzkumným záměrem MSM 0021622404
Úvod
Koncept default mode sítě (z anglického default mode network – DMN, kde označení default mode představuje „bazální“ mozkovou aktivitu) se začal rozvíjet v 90. letech minulého století, kdy za pomocí neurozobrazovacích metod (nejdříve studie prostřednictvím pozitronové emisní tomografie – PET, později i fMR) byly v průběhu provádění cílených úkolů zaznamenány poklesy aktivity (tzv. deaktivace) v určitých oblastech mozku, zatímco jiné oblasti mozku, přímo zodpovědné za reakce na tyto podněty, se aktivovaly. Stejné oblasti deaktivující se v průběhu úkolu naopak vykazovaly určitou aktivitu za stavu klidu či pasivní oční fixace, kdy žádný úkol prováděn nebyl [1]. Signál, kterým se tato aktivita za klidu projevovala, měl typický průběh a byl označen jako nízkofrekvenční fluktuace. Tyto fluktuace o frekvenci do 0,1 Hz vykazovaly nápadné shody u jednotlivých vyšetřovaných skupin a pravděpodobně představují spontánní fluktuující neuronální aktivitu, často označovanou také jako „resting state“ (klidový stav) [1].
Regionální deaktivace měřitelné pomocí fMR v průběhu provádění cílených úkolů pravděpodobně představují mozkovou aktivitu, která je nepřetržitá v průběhu klidového stavu a snižována v průběhu cílených reakcí. Odtud plyne i název „default mode“ funkce mozku. Zjistilo se, že některé oblasti vykazující takovou aktivitu jsou organizovány do určitého systému a byly označeny jako „default mode“ síť. Součásti této sítě jsou zadní cingulum/přilehlý praecuneus, přední cingulum/ventromediální prefrontální kortex, gyrus angularis/lobulus parietalis inferior, dále též mediální temporální oblasti včetně hipokampu.
Nicméně pravděpodobně všechny oblasti mozku mohou vykazovat určitý stupeň organizované „default“ aktivity [1]. Byly již identifikovány další mozkové oblasti, které tyto nízkofrekvenční fluktuace ve stavu klidu vykazují, například zadní parietální, frontotemporální a přední temporální oblasti, limbický lalok, ale také podkorové oblasti, jako thalamus, bazální ganglia či mozeček [2,3]. Na rozdíl od DMN se v průběhu kognitivního úkolu tyto oblasti aktivují. Představují tak tzv. anti-korelující aktivitu k deaktivacím v DMN a obě tyto komponenty mohou být součástí jedné sítě, přičemž aktivující se součást by mohla zajišťovat reakce na neočekávané či nové události [4].
Funkce DMN zatím nebyla objasněna, na vysvětlení jejího významu bylo vysloveno několik hypotéz. Lze předpokládat, že mozkové reakce se neobjevují jen jako pouhá odpověď na momentální požadavky prostředí, ale že mají i určitou vnitřní aktivitu stojící na pozadí těchto odpovědí. Tento předpoklad podporuje skutečnost, že pouze 0,5–1 % celkové energie mozku je vynaloženo na procesy související s reakcemi na podněty z prostředí; 60–80 % energie je využito na podporu komunikace mezi neurony [1]. Přítomnost DMN může být odrazem této vnitřní aktivity mozku a v rámci jejích možných funkcí, zatím ne zcela objasněných, může DMN hrát roli v interpretaci a předvídání stimulů z prostředí či udržování „rovnováhy“ mezi excitačními a inhibičními vstupy a tím ulehčovat odpovědi na podněty. Předpokládá se rovněž funkce DMN v kognitivních procesech [1,5].
Cílem naší práce bylo zjistit, které z možných metod zpracování získaných fMR dat nám nejlépe umožní identifikovat DMN a reprodukovat výsledky publikované v literatuře. Dalším naším cílem bylo ověřit hypotézu, že míra úspěšnosti v kognitivních testech pozitivně koreluje s mírou deaktivací v hlavních oblastech zapojených do DMN.
Metodika
Do naší pilotní studie bylo zařazeno 10 zdravých dobrovolníků (sedm žen, tři muži), všichni praváci a všichni bez onemocnění mozku, demence či psychiatrického onemocnění, průměrný věk 61,7 ± 6,7 let. Všichni dobrovolníci podstoupili vyšetření kognitivních funkcí Addenbrookským kognitivním testem [6]. Vyšetření pomocí fMR sestávalo ze tří částí: a) vizuální kognitivní úkol, b) klidový stav neboli „resting state“, c) anatomické snímky.
Addenbrookský kognitivní test (Addenbrook cognitive examination, ACE)
Slouží k detailnější diagnostice demence. Test sestává z 18 úkolů uspořádaných do pěti domén – hodnotí se pozornost a orientace, paměť, verbální fluence, jazyk a zrakově-prostorové schopnosti. Jeho součástí je rovněž MMSE (Mini mental state examination). Maximální počet bodů dosažitelný v tomto testu je 100, minimální počet je 0 bodů. Skóruje-li pacient ≤ 88 bodů, je senzitivita pro diagnostiku demence 94 % a specificita 89 % [6].
Klidový stav
Před zahájením snímání klidových dat byli dobrovolníci vyzváni, aby leželi v klidu se zavřenýma očima a snažili se odpočívat, nehýbat se a na nic konkrétního nemyslet (nic konkrétního v duchu neřešit). Snímání trvalo 15 minut.
Kognitivní vizuální úkol
Úloha byla navržena podle úkolu použitého ve studii dle Rabina et al [7] a spočívala v testování vizuální paměti. Zkoumané osoby měly za úkol sledovat obrázky s různou tematikou promítané v zrcadle hlavové cívky a snažit se je zapamatovat. Celkový počet obrázků byl 60, obrázky byly promítány v sekvencích po deseti, přerušených deseti kontrolními obrazci. Vyšetření začínalo i končilo sekvencí deseti kontrolních obrazců (kontrolních obrazců bylo tedy celkem 70). Délka promítání obrazců byla 3,5 s; délka pauzy mezi obrazci 550 ms (obr. 1).

fMR vyšetření
Akvizice byla provedena na MR tomografu Siemens Magnetom Symphony 1,5T. Při kognitivním úkolu byla naměřena série 260 objemů (skenů) T2* vážených snímků pomocí echoplanárního zobrazování (EPI). Každý sken sestával z 20 transverzálních řezů s rozlišením 64 × 64 voxelů, zorným polem (field of view, FOV) 240 × 240 mm a z toho vyplývající velikostí pixelu 3,75 × 3,75 mm. Tloušťka řezu byla 5 mm. Repetiční čas akvizice byl 2 050 ms, čas TE byl 50 ms a sklápěcí úhel 90°. Pro klidová data byla naměřena série 300 skenů T2* vážených EPI snímků. Každý sken sestával z 32 transverzálních řezů s rozlišením 64 × 64 voxelů, FOV 220 × 220 mm a z toho vyplývající velikostí pixelu 3,4375 × 3,4375 mm. Tloušťka řezu byla 3,5 mm. Repetiční čas akvizice byl 3 000 ms, čas TE byl 40 ms a sklápěcí úhel 90°. Po sériích funkčních snímků následovala akvizice T1 vážených podrobných anatomických snímků mozku o 160 sagitálních řezech s tloušťkou 1,18 mm a rozlišením 256 × 256 voxelů převzorkovaných na 512 × 512.
Po naměření fMR dat následovalo testování rekognice (znovupoznání). Subjektům bylo v časovém odstupu cca 30 minut od fMR vyšetření promítáno na počítačovém monitoru všech 60 obrázků viděných v průběhu fMR vyšetření náhodně promíchaných se 60 novými obrázky (celkem tedy 120 obrázků). Pod obrázky se nacházela tlačítka pro označení známého či nového obrázku a tlačítko „pauza“. Rychlost střídání obrázků v průběhu testování rekognice byla stejná jako v průběhu fMR vyšetření. Podle toho, zda subjekt obrázek rozpoznal jako již viděný v průběhu fMR vyšetření, bylo zmáčknuto tlačítko „známý“, pokud tento obrázek subjekt rozpoznal jako neznámý, bylo zmáčknuto tlačítko „nový“. V některých případech, když subjekt nestihl reagovat či obrázek nebyl schopen zařadit, zůstal obrázek neoznačen. Pro případ nutnosti dočasně přerušit testování sloužilo tlačítko „pauza“. Program byl ovládán pomocí myši a podle toho, zda subjekt zvládal práci s myší, pracoval s programem subjekt (ovládání myši pravou rukou), v opačném případě bylo testování zprostředkováno vyšetřovatelem.
Analýza fMR dat
Data z kognitivního úkolu byla zpracována pomocí programu SPM5 (http://www.fil.ion.ucl.ac.uk/spm/) v prostředí Matlab 7.5. Funkční snímky byly korigovány na zjištěný pohyb, normalizovány do standardního stereotaktického prostoru pomocí MNI šablony a prostorově vyhlazeny Gaussovým filtrem s FWHM = 8 mm. Statistické zpracování bylo provedeno pomocí obecného lineárního modelu implementovaného v programu SPM5. Z dat byly odfiltrovány nízké frekvence (cut-off = 128 s) a byla provedena korekce na autokorelaci signálu. Regresor zájmu byl modelován konvolucí stimulační funkce s průběhem hemodynamické odezvy. Dále byly použity regresory představující odhadnutý průběh pohybu (šest regresorů, tři pro posuny a tři pro rotace), které sloužily k eliminaci pohybových artefaktů ve výsledných statistických parametrických mapách. Výsledek pak byl zobrazen pomocí t-statistiky, kdy bylo sledováno významné zvýšení nebo naopak snížení (tzv. deaktivace) fMR signálu ve shodě s průběhem stimulace. Následně byla provedena korelační analýza (tzv. seed funkční konektivita) vycházející z oblasti zájmu lokalizované v zadním cingulu/precuneu, určené při zobrazení deaktivace u jednotlivých subjektů. Ta spočívala v identifikaci oblastí, které měly podobný průběh signálu jako ve vybraných oblastech zájmu. Byla provedena skupinová analýza metodou náhodných efektů (random effect analysis) pro zobrazení aktivace a deaktivace dle kognitivního úkolu a pro korelační seed konektivitu. Při skupinové analýze byl jako kovariát použit věk subjektů. K prohlížení výsledných statistických parametrických map byla použita hladina významnosti p < 0,0001 a minimální rozsah aktivace 5 voxelů.
Dále jsme sledovali, jak fMR signál v průběhu deaktivací koreluje s výkonem v kognitivních testech, konkrétně s úspěšností ve vizuálním paměťovém úkolu, Addenbrookském kognitivním testu a jeho dvou subtestech – testu verbální fluence a testu paměti.
Klidová data byla zpracována pomocí ICA v programu GIFT verze 2.0a (http://icatb.sourceforge.net/) v prostředí Matlab 7.5. U každé osoby byl identifikován optimální počet komponent a ten použit při individuálním zpracování. Komponenta představující „resting state“ byla identifikována na základě prostorové korelace s maskou obsahující oblasti zapojené do DMN. Tato maska je součástí programu GIFT.
Pro ohodnocení podobnosti jednotlivých přístupů zobrazení DMN byly vypočteny korelační koeficienty mezi mapami deaktivací a ICA komponentami a mezi mapami seed konektivity a ICA komponentami. K prohlížení zpracovaných dat byl použit program Xjview rovněž v prostředí Matlab 7.5.
Výsledky
Behaviorální data
V naší studii jsme pro testování kognitivních funkcí použili jednak Addenbrookský kognitivní test (a), jednak kognitivní vizuální úkol v průběhu fMR vyšetření s následným testováním rekognice (b).
ad a) Addenbrookský kognitivní test
Průměrná úspěšnost v ACE byla 94,7 ± 3,8 bodů, žádný z dobrovolníků neměl počet bodů menší než 89 (cut-off skóre pro demenci je 88). Detailnější výsledky všech subtestů viz tab. 1. Pro korelační analýzu bylo použito celkové skóre a dále subskóre při testu verbální fluence a paměti, a to proto, že v těchto doménách vykazovali naši dobrovolníci největší variabilitu (tab. 1).

ad b) Testování rekognice
Průměrná úspěšnost v paměťovém vizuálním úkolu byla 73,2 ± 7,3 %, tj. dobrovolníci průměrně rozpoznali správně 87,8 ± 8,1 obrázků z celkového množství 120 obrázků jako známých či neznámých. Výsledky u jednotlivých subjektů viz tab. 2. Počty správně rozpoznaných obrázků u jednotlivých subjektů jsme rovněž použili pro následnou korelační analýzu.
Sledování default-mode sítě
Pro sledování DMN jsme použili (a) klidová data a zpracování pomocí ICA (b) hodnocení prosté „deaktivace“ v průběhu kognitivního vizuálního úkolu, (c) hodnocení ad b) s použitím „seed“ funkční konektivity vycházející z oblasti zájmu zadní cingulum(PCC)/praecuneus.
Ad a) Klidová data
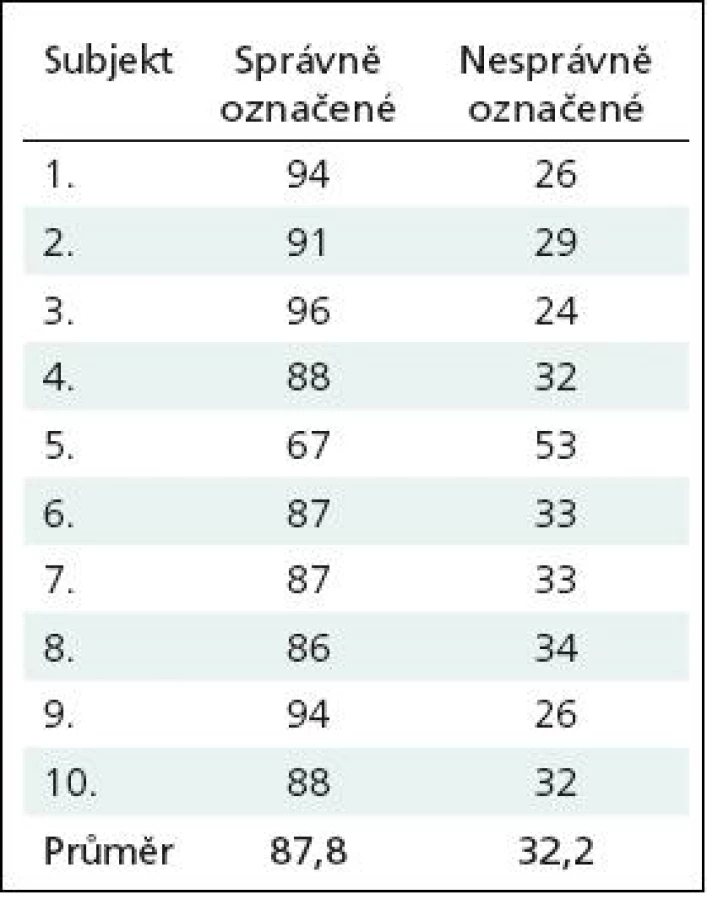
Obr. 2 demonstruje výsledek ICA analýzy ve vybrané komponentě, která nejlépe korelovala s maskou představující DMN, zobrazené v programu Xjview. Aktivní oblasti v průběhu klidového stavu zde viditelné jsou zejména PCC/precuneus, ventromediální prefrontální kortex (VMPFC)/přední cingulum (ACC) a gyrus angularis (GA)/lobulus parietalis inteferior (LPI).
Ad b) Zpracování pomocí ICA
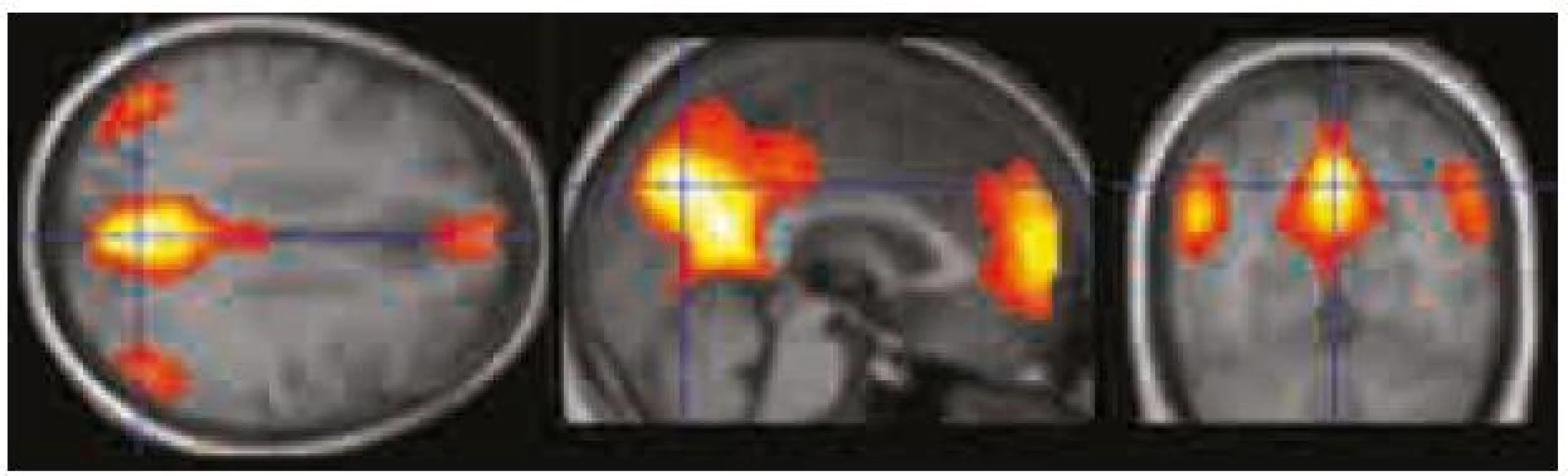
Obr. 3 představuje oblasti deaktivací v průběhu vizuálního úkolu zobrazené pomocí programu Xjview. I zde dochází ke změnám MR signálu v obdobných oblastech jako při hodnocení metodou a), nicméně rozsah a signifikance statistické mapy modelující DMN je mnohem nižší. Obraz komponenty představující DMN v ICA analýze v tomto případě koreluje s obrazem prosté deaktivace v průběhu kognitivního úkolu s korelačním koeficientem r = 0,09.
Ad c) Hodnocení s použitím „seed“ funkční konektivity
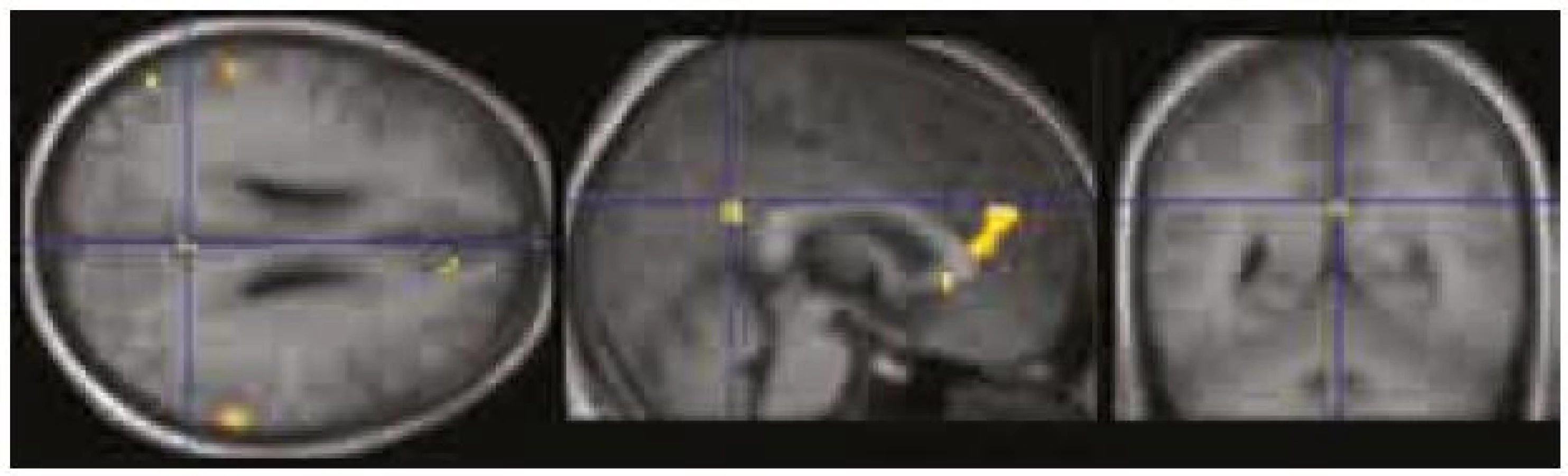
Obr. 4 ukazuje výsledky „seed analýzy“ vycházející z oblasti zájmu PCC/precuneus (v programu Xjview). V porovnání s výsledky prosté deaktivace (obr. 3) je statistická mapa oblastí získaná na podkladě „seed analýzy“ rozsáhlejší a lépe koreluje s výsledky ICA analýzy (r = 0,29).
Korelace s behaviorálními daty
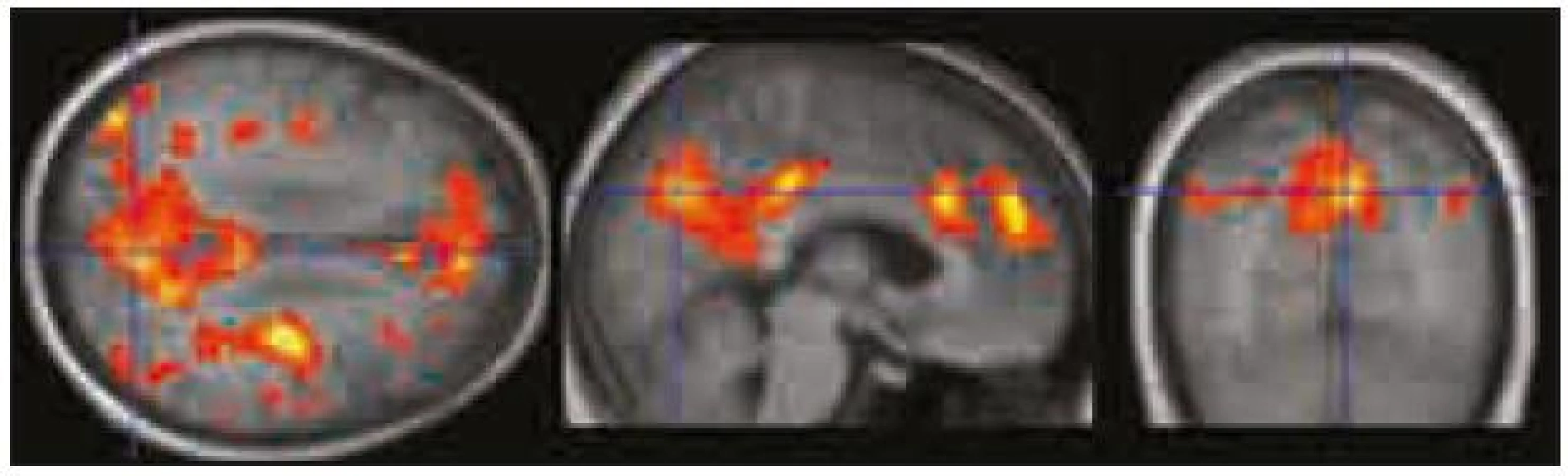
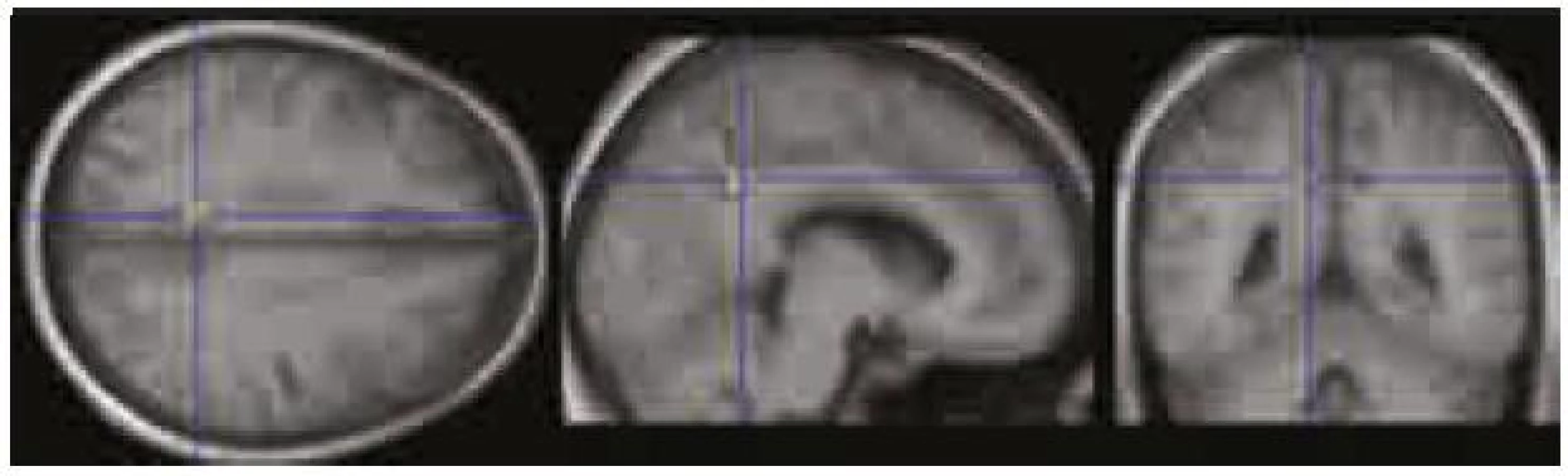
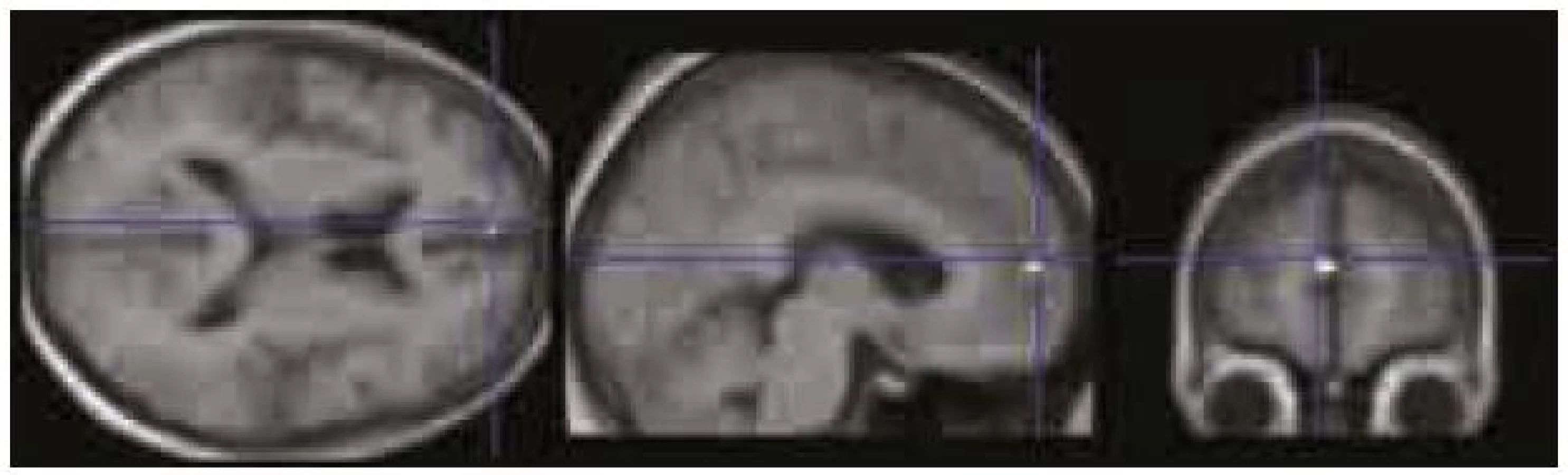
Použili jsme výsledky úspěšnosti rekognice ve vizuálním paměťovém úkolu (tab. 2) a celkové skóre v ACE a jeho dvou subtestech – testu verbální fluence a testu paměti (tab. 1). Našli jsme signifikantní zápornou korelaci (p < 0,0001 nekorigovaně) mezi MR signálem v PCC/precuneu a výkonem v testu verbální fluence, tj. čím vyšší byl výkon v testu verbální fluence, tím významnější byla deaktivace v PCC/precuneu (obr. 5). Další záporná korelace, ale na nižší hladině významnosti (p < 0,001 nekorigovaně) byla zjištěna mezi MR signálem v oblasti VMPFC a skórem rekognice ve vizuálním paměťovém úkolu, tj. čím lepší byla rekognice, tím významnější byla deaktivace ve VMPFC (obr. 6).
Diskuze
V naší pilotní studii jsme ověřili schopnost detekovat DMN u zdravých dobrovolníků pomocí tří různých přístupů, a to pomocí ICA analýzy, pomocí zobrazení prosté deaktivace průběhu kognitivního úkolu a pomocí funkční seed konektivity vycházející z dat získaných při hodnocení prosté deaktivace v průběhu kognitivního paměťového úkolu. Pro zobrazení funkční seed konektivity jsme jako oblast zájmu (seed) zvolili oblast zasahující do zadního cingula nebo precuneu. Oblast zadního cingula a precuneu byla použita pro tento účel i v některých předchozích obdobných studiích [5,8]. Bez ohledu na použitou metodu jsme byli schopni detekovat tři oblasti zapojené do DMN, a to: zadní cingulum/precuneus, ventromediální prefrontální kortex/přední cingulum a gyrus angularis/lobulus parietalis inferior. Naše výsledky jsou tedy ve shodě s oblastmi popsanými v literatuře [1,5,8–13].
Nejlepší zobrazení DMN přineslo zpracování pomocí ICA analýzy. Výsledky ICA analýzy jsme proto dále porovnávali s prostými deaktivacemi a s výsledky funkční seed konektivity v průběhu kognitivního úkolu. Zjistili jsme, že výsledky ICA analýzy významně lépe korelují s výsledky seed konektivity ze zadního cingula/precuneu než s prostým obrazem deaktivace (viz též obr. 2–4).
Výsledky funkčních neurozobrazovacích metod nasvědčují tomu, že precuneus má reciproční spoje se zadním i předním cingulem, retrosplenickým kortexem, zadní premotorickou a suplementární motorickou areou. Obecně má precuneus rozsáhlá spojení s vyššími korovými a podkorovými asociačními oblastmi, přičemž nebyly pozorovány přímé spoje s primárními senzorickými oblastmi. Obecně je precuneus zapojen spíše do rozsáhlých procesů hodnotících vysoce integrované a asociativní informace než do přímého zpracovávání vnějších podnětů [14]. Precuneus hraje roli ve vybavování epizodické paměti, zrakově-prostorové představivosti, zpracování informací týkajících se vlastního já a v udržování bdělého stavu vědomí [14].
Mediální prefrontální kortex má spoje s limbickými oblastmi důležitými pro paměť, motivaci a afektivitu, je součástí mnoha kognitivních funkcí zahrnujících behaviorální inhibici, pozornost a plánování, ukládání a vybavování epizodické paměti [15–18]. Důležitá je jeho role v procesech souvisejících s vlastním já, jako sebehodnocení, rozhodování či přerozdělování pozornosti mezi informace související s vlastním já nebo s vnějšími podněty [16]. Obecně se dá říct, že mediální prefrontální kortex je spíše spojen s procesy zaměřenými na vlastní osobu jako takovou, zatímco zadní cingulum spíše na vlastní já v kontextu sociálním [19].
Avšak jen málo je známo o behaviorálním korelátu deaktivací v těchto oblastech (či v posteromediálních parietálních a mediálních prefrontálních oblastech obecně). Jednou z možností je, že v době, kdy je jednotlivec při vědomí, ale nevykonává žádný kognitivní úkol (tj. je ve stavu klidu), precuneus a další propojené oblasti, jako zadní cingulum a ventromediální prefrontální kortex, provádějí třídění informací týkajících se vlastního já a vnějšího prostředí, účastní se procesů spojených s konzolidací epizodické paměti a jejího vybavování, zpracovávání informací ve formě spontánních mimovolně plynoucích myšlenek a mentálních představ a manipulací s těmito informacemi za účelem řešení problémů a plánování [9]. Když je pak klidový stav přerušen kognitivním procesem, dochází k přerozdělení zdrojů využívaných pro třídění a manipulaci s „vnitřními“ informacemi ve prospěch aktivity spojené s koncentrovanou volní odpovědí na zevní stimuly a nutné pro úspěšné vykonání kognitivního úkolu [14].
V naší studii jsme zjistili, že čím vyšší byla úspěšnost v testu verbální fluence, tím výraznější byla deaktivace v oblasti zadního cingula/precuneu. V této oblasti Schlösser et al 1998 [20] také pozorovali (pomocí fMR) snížení BOLD signálu přímo v průběhu provádění testu verbální fluence. Při zkoumání korelace deaktivací v DMN s výsledky vizuálního paměťového úkolu při použité hladině významnosti p < 0,0001 nekorigovaně nebyl patrný žádný signifikantní výsledek, avšak při snížení hladiny na p < 0,001 se objevila záporná korelace mezi mírou deaktivace v oblasti ventromediálního prefrontálního kortexu a výkonem v paměťovém vizuálním úkolu (testování rekognice). Tento námi zjištěný trend bude ovšem nutné ověřit studiemi na větším souboru subjektů.
Ačkoliv se jednalo o vyšetření zdravých dobrovolníků s normálním kognitivním výkonem dle skríningového ACE-R testu, zjistili jsme, že kognitivní výkon přinejmenším v subtestu verbální fluence souvisel s mírou deaktivace v oblasti zadního cingula/precuneu, tj. v oblasti zapojené do DMN. To by podporovalo hypotézu o významu DMN pro kognitivní procesy v mozku. Tato hypotéza je ve shodě i s výsledky studií zaměřených na změnu DMN u pacientů s Alzheimerovou nemocí či mírným kognitivním deficitem (Mild cognitive impairment, MCI), kde byla zjištěna snížená resting-state aktivita či konektivita mezi jednotlivými oblastmi DMN [8,10,11,13,16].
Proces přechodu z bazální resting state aktivity do stavu deaktivací v průběhu provádění kognitivního úkolu je však proces složitější, ovlivněný aktivitou četných dalších nervových sítí. Proto snížení resting state aktivity nemusí vždy paušálně předpokládat i snížení míry deaktivací a naopak. DMN tvoří s dalšími oblastmi mozku funkčně propojený systém, jehož správná funkce závisí na adekvátním přerozdělování („switch“) aktivity mezi DMN a jinými sítěmi. Teoreticky tak narušení deaktivací může být výsledkem „chybného“ přerozdělení této aktivity spojeného s nedostatečnou funkcí některého z těchto zapojených systémů, přičemž samotný resting state narušen být nemusí. Nebo může docházet ke kompenzatorním přesunům aktivací/deaktivací ve prospěch DMN či jiných sítí. Výsledky hodnocení DMN pomocí ICA analýzy z resting state dat mohou mít tedy jiný patofyziologický podklad než hodnocení DMN pomocí deaktivací naměřených v průběhu kognitivního úkolu. ICA je navíc zatížena problémem správného výběru komponenty.
Výsledky naší pilotní studie je nutno ověřit na větší skupině zdravých dobrovolníků. Dalším krokem bude porovnání se skupinou věkově vázaných pacientů s Parkinsonovou nemocí, Alzheimerovou demencí a demencí u Parkinsonovy nemoci. Je možné, že vyšetření DMN by mohlo v budoucnu sloužit jako časný marker kognitivní poruchy a demence [8,10,11,13].
Závěr
Naše metoda je vhodná pro studium DMN, kterou jsme pozorovali s pomocí fMR jednak za klidového stavu (jako tzv. resting state), jednak v průběhu provádění kognitivního úkolu (jako tzv. deaktivace). Míra těchto deaktivací koreluje s kognitivním výkonem dobrovolníků. Je možné, že vyšetření DMN by mohlo sloužit jako časný marker kognitivní poruchy a/nebo demence. Výsledky naší studie jsou ovšem pilotní a je nutno je ověřit na větší skupině zdravých subjektů.
doc. MUDr. Irena Rektorová, Ph.D.
I. neurologická klinika
LF MU a FN u sv. Anny
Pekařská 53
656 91 Brno
e-mail: irena.rektorova@fnusa.cz
Přijato k recenzi: 24. 11. 2009
Přijato do tisku: 20. 4. 2010
Zdroje
1. Raichle ME, Snyder ZE. A default mode of brain function: a brief history of an evolving idea. Neuroimage 2007; 37(4): 1083–1090.
2. Robinson S, Basso G, Soldati N, Sailer JJ, Bruzzone L, Krypsin-Exner I et al. A resting state network in the motor control circuit of the basal ganglia. BMC Neurosci 2009; 10: 137.
3. Calhoun VD, Kiehl KA, Pearlson GD. Modulation of temporally coherent brain network estimated using ICA at rest and during cognitive tasks. Hum Brain Mapp 2008; 29(7): 828–838.
4. Broyd SJ, Demanuele C, Debener S, Helps SK, James CJ, Sonuga-Barke EJ. Default-mode brain dysfunction in mental disorders: a systematic review. Neurosci Biobehav Rev 2009; 33(3): 279–296.
5. Greicius MD, Krasnow B, Reiss AL, Menon V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proc Natl Acad Sci U S A 2003; 100(1): 253–258.
6. Hummelová-Fanfrdlová Z, Rektorová I, Sheardová K, Bartoš A, Línek V, Ressner P et al. Česká adaptace Addenbrookského kognitivního testu. Československá psychologie 2009; 53: 376–388.
7. Rabin ML, Narayan VM, Kimberg DY, Casasanto DJ, Glosser G, Tracy JI et al. Functional MRI predicts post-surgical memory following temporal lobectomy. Brain 2004; 127(10): 2286–2298.
8. Greicius MD, Srivastava G, Reiss AL, Menon V. Default-mode network activity distinguishes Alzheimer´s disease from healthy aging: Evidence from functional MRI. Proc Natl Acad Sci U S A 2004; 101(13): 4637–4642.
9. Gusnard DA, Raichle ME. Searching for a baseline: functional imaging and the resting human brain. Nat Rev Neurosci 2001; 2(10): 685–694.
10. Wang L, Zang Y, He Y, Liang M, Zhang X, Tian L et al. Changes in hippocampal connectivity in the early stages of Alzheimer’s disease: evidence from resting state fMRI. Neuroimage 2006; 31(2): 496–504.
11. Rombouts SA, Barkhof F, Goekoop R, Stam CJ, Scheltens P. Altered resting state network in mild cognitive impairment and mild Alzheimer’s disease: an fMRI study. Hum Brain Mapp 2005; 26(4): 231–239.
12. Harrison BJ, Pujol J, López-Solà M, Hernández-Ribas R, Deus J, Ortiz H et al. Consistency and functional specialization in the default mode brain network. Proc Natl Acad Sci U S A 2008; 105(28): 9781–9786.
13. Supekar K, Menon V, Rubin D, Musen M, Greicius MD. Network analysis of intrinsic functional brain connectivity in Alzheimer’s disease. PLoS Comput Biol 2008; 4(6): e1000100.
14. Cavanna AE, Trimble MR. The precuneus: a review of its functional anatomy and behavioral correlates. Brain 2006; 129(3): 564–583.
15. Briand LA, Gritton H, Howe WM, Zoung DA, Sarter M. Modulators in concert for cognition: Modulator interactions in the prefrontal cortex. Prog Neurobiol 2007; 83(2): 69–91.
16. Dumontheil I, Burgess PW, Blakemore SJ. Development of rostral prefrontal cortex and cognitive and behavioural disorders. Dev Med Child Neurol 2008; 50(3): 168–181.
17. Lepage M, Ghaffar O, Nyberg L, Tulving E. Prefrontal cortex and episodic memory retrieval mode. Proc Natl Acad Sci U S A 2000; 97(1): 506–511.
18. Ranganath C, Johnson MK, D’Esposito M. Left anterior prefrontal activation increases with demands to recall specific perceptual information. J Neurosci 2000; 20(22): 108.
19. Johnson MK, Raye CL, Mitchell KJ, Touryan SR, Greene EJ, Nolen-Hoeksema S. Dissociating medial frontal and posterior cingulate activity during self-reflection. Soc Cogn Affect Neurosci 2006; 1(1): 56–64.
20. Schlösser R, Hutchinson M, Joseffer S, Rusinek H, Saarimaki A, Stevenson J et al. Functional magnetic resonance imaging of human brain activity in a verbal fluency task. J Neurol Neurosurg Psychiatry 1998; 64(4): 492–498.
Štítky
Dětská neurologie Neurochirurgie NeurologieČlánek vyšel v časopise
Česká a slovenská neurologie a neurochirurgie

2010 Číslo 5
Nejčtenější v tomto čísle
- Neuralgie nervus pudendalis – kazuistika
- Vývoj technik PLIF a TLIF
- Syndrom útlaku ulnárního nervu v oblasti lokte – přehled operačních technik a srovnání jejich výsledků
- Střelná poranění hlavy a mozku