
Na dosah dolnímu fronto-okcipitalnímu fasciculu s pomocí disekce dle Klinglera a DTI traktografie
Reaching the inferior fronto-occipital fascicle with the help of Klingler dissection and DTI tractography
Aim: The aim of this study is to provide both image sources and a verbal description to allow the mental visualization of the course of the inferior fronto-occipital fascicle (IFOF) when looking at the brain from the lateral side, and to highlight the clinical importance of this associative white matter tract. Methods: In the three hemispheres of the brains of deceased donors, the IFOF was dissected using the Klingler method, with the aim to preserve as many intact cortical structures as possible. The spared cortical structures of the brain are good reference points for orientation on the brain surface. Diffusion tensor imaging (DTI) tractography was performed as another method to study the anatomical course of the IFOF. The data for the tractography were obtained using MRI examination in a healthy volunteer. Results: It was possible to dissect the IFOF in all three hemispheres. The course of the IFOF was documented in photographs of the dissections. Similarly, the course of the IFOF is depicted through the use of tractographic reconstructions and projections of these reconstructions in the MRI image of the brain. Both methods provide very similar results when it comes to IFOF anatomy. Conclusion: The availability of neuronavigation or other technological instruments did not reduce the need for knowledge of anatomy. The authors hope that the results presented in this project can serve to expand one’s knowledge or at least to awaken an interest in anatomy.
Keywords:
human brain anatomy – white matter – fiber dissection – Diffusion tensor imaging – Tractography – inferior fronto-occipital fasciculus – semantic system
Authors:
E. Neuman 1; O. Šandor 2; M. Hána 2; M. Keřkovský 3; M. Joukal 2; M. Smrčka 1
Authors‘ workplace:
Neurochirurgická klinika FN Brno
1; Anatomický ústav, LF MU Brno
2; Klinika radiologie a nukleární, medicíny FN Brno
3
Published in:
Cesk Slov Neurol N 2021; 84/117(2): 139-150
Category:
Original Paper
doi:
https://doi.org/10.48095/cccsnn2021139
Overview
Cíl: Cílem této studie je poskytnout obrazové podklady i slovní popis k umožnění představy anatomického průběhu fasciculus fronto-occipitalis inferior (inferior fronto-occipital fascicle; IFOF) při pohledu na mozek z laterální strany a také poukázat na klinickou důležitost této asociační dráhy bílé hmoty. Metoda: Ve třech hemisférách mozků zemřelých dárců byl Klinglerovou metodou vypreparován IFOF, a to se snahou ponechat co možná nejvíce původních kortikálních struktur. Ponechané kortikální struktury mozku umožňují udržení orientace na povrchu mozkové hemisféry. Jako další metoda ke studiu anatomického průběhu IFOF bylo provedeno zobrazení tenzorů difuze (diffusion tensor imaging; DTI) traktografie z dat získaných vyšetřením MR zdravého dobrovolníka. Výsledky: IFOF se ve všech třech hemisférách podařilo vypreparovat. Průběh IFOF byl zdokumentován na fotografiích preparátů. Podobně je průběh IFOF znázorněn pomocí traktografických rekonstrukcí a na projekcích výsledků traktografií do MR obrazu mozku. Obě metody poskytují, pokud jde o anatomii IFOF, velmi podobné výsledky. Závěr: Dostupnost neuronavigace a případně jiných technologických vymožeností nijak nesnížila potřebu znalosti anatomie. Autoři doufají, že výsledky předkládané v práci mohou posloužit k rozšíření znalostí nebo alespoň probudit zájem o anatomii.
Klíčová slova:
anatomie lidského mozku – bílá hmota – disekce nervových drah – zobrazení tenzorů difuze – traktografie – fasciculus fronto - occipitalis inferior – sémantický systém
Úvod
Ještě v ne úplně vzdálené době, před 10–15 lety, se při plánování a provádění neurochirurgických operací braly v úvahu především funkčně důležité kortikální (tzv. elokventní) oblasti, zatímco o nervových dráhách (fasciklech nervových vláken) se v kontextu možného porušení příliš neuvažovalo. Tomuto přístupu odpovídal teoretický předpoklad, že každá funkce má na kůře mozku jedno určité místo, které se nesmí porušit. V dalších letech se ale pohled na způsob fungování mozku změnil. Místo existence statických, pevně daných funkčních kortikálních oblastí se ukázalo, že činnost mozku probíhá na podkladě mnoha hierarchicky uspořádaných, vzájemně se ovlivňujících a funkčně se překrývajících neuronálních sítí [1].
Důležitost mozkové kůry jakožto oblasti obsahující těla neuronů samozřejmě zůstává, ale pokud jde o funkci, pozornost bylo potřeba rozšířit na jejich rozsáhlé spoje. Je již dlouho známo, že léze bílé hmoty jsou prognosticky horší než léze kortikální, a to ze zkušeností s CMP. „Přerušení kabelu“ mívá horší následky než zničení kortikální oblasti, neboť vede k funkčnímu odpojení širokých navazujících oblastí. Na druhou stranu má toto „síťové“ uspořádání s možným (novým a dodatečným) zapojením mnoha vzdálených oblastí za následek existenci určité schopnosti regenerace poškozených funkcí, což je nazýváno plasticitou [2].
Platí obecné pravidlo, že čím vzdálenější je dráha od povrchu mozku, tím menší plasticitu lze očekávat, neboť na hlouběji uložené spoje jsou přes povrchově uložené spoje navázány široké kortikální oblasti – vlákna se vějířovitě sbíhají od povrchu a směrem do hloubky se koncentrují. To v neurochirurgické praxi znamená, že hluboko uložené dráhy jsou hranicemi proveditelnosti resekcí a mohou limitovat dosažitelnost lézí [3].
Disekce vláken bílé hmoty je starou neuroanatomickou technikou používanou již od 17. století. Metodou disekce bílé hmoty se zabývali anatomové jako Thomas Willis (1621–1675) a Raymond Vieussens (1635–1717). Význačné místo mezi mnohými, jež se v minulosti zabývali disekcí nervových drah, patří Josefu Klinglerovi (1888–1963). Klingler díky svému objevu, že přechodné zmrazení formalínem fixovaného mozkového preparátu vede k rozvolnění svazků vláken, a díky pozoruhodnému úsilí, které věnoval preparaci vláken bílé hmoty, vytvořil dílo, které je dodnes cenným zdrojem informací o struktuře bílé hmoty mozku [4].
Jednou z nejzajímavějších asociačních drah bílé hmoty je fasciculus fronto-occipitalis inferior (inferior fronto-occipital fascicle; IFOF). Už jeho objevování má zajímavou historii. První zmínku o vláknech spojujících okcipitální lalok s frontálním učinil německý anatom Karl Burdach v roce 1822. Při studiu fasciculus longitudinalis inferior (který spojuje okcipitální lalok s temporálním lalokem při bazi temporálního laloku) zjistil, že část vláken, která považoval za vlákna patřící k fasciculus longitudinalis inferior, se odděluje, stáčí se do inzuly a pokračují dále do frontálního laloku [5]. Burdachův popis těchto vláken souhlasí velmi dobře s tím, co se dnes nazývá IFOF.
Francouzský anatom Jules Dejerin v roce 1895 zavedl termín „fasciculus fronto-occipitalis“ pro spoje, které pozoroval mezi frontálním a okcipitálním lalokem a které považoval je za samostatnou anatomickou jednotku [5]. Avšak centrální část tohoto svazku vyobrazoval (chybně) nad nucleus lentiformis, zatímco IFOF vede po předním a dolním okraji tohoto jádra. A tak v roce 1909 australsko-irský student medicíny Edward Curran ani poté, co provedl disekci 200 preparátů mozku, nebyl schopen tento Dejerinem popisovaný fasciculus fronto-occipitalis nalézt. Nalezl však jiné spojení mezi frontálním a okcipitálním lalokem, ale jelikož si nejspíše netroufal zpochybnit nález autority (tedy Dejerina), nazval jej odlišně – jako fasciculus fronto-occipitalis inferior [6].
Zajímavé také je, že Curran uvádí IFOF slovy jako „nový a dosud nepopsaný svazek vláken“ [6]. Přitom ale již v roce 1906 francouzský anatom Jean Baptiste P. Trolard vydal práci „Le faisceau longitudinal inférieur du cerveau“ [5] s popisem a nákresy přesně zachycujícími průběh fasciklů vláken odpovídajících IFOF. Curran ale svůj objev učinil pravděpodobně bez znalostí prací Trolarda a Burdacha.
V dalších 100 letech se pak IFOF ve vědeckých pracích objevoval jen sporadicky, jako by zmizel ze zájmu badatelů. V 50. letech minulého století jej ale ve své práci zobrazuje Josef Klingler [4].
Po vypracování metody zobrazení tenzorů difuze (difussion tensor imaging; DTI) traktografie se zájem o IFOF náhle obnovil. Jako první se o něm v této souvislosti zmiňuje Catani v roce 2002, který vypracoval atlas drah bílé hmoty založený na DTI traktografii [7]. A od té doby stále narůstají poznatky o funkčním významu tohoto svazku, získané ponejvíce z pozorování při awake kraniotomiích.
Ještě do nedávné doby se ale objevovaly vážné pochybnosti, zda IFOF doopravdy existuje a zda se nejedná o chybnou interpretaci dat. Pochybnosti vznikaly proto, že v mozcích zvířat (konkrétně makaků, čeleď kočkodanovití) se IFOF nedařilo nalézt [8]. Vzhledem k tomu, že se IFOF u člověka naopak dařilo zobrazit pomocí traktografie a i při disekci, spekulovalo se, že by IFOF mohl být jedinečným svazkem existujícím pouze u člověka [9]. Tuto domněnku umocňovalo zejména to, že IFOF zprostředkovává schopnosti typické pro člověka, jako jsou řeč, schopnost zobecňovat a pracovat s koncepty. Současně však z evolučního hlediska byla tato možnost nepravděpodobná, neboť vývojově starší dráhy jsou vždy uloženy hlouběji a bývají dlouhé, což IFOF charakterizuje. Tento rozpor vyřešil až italský neurochirurg Silvio Sarrubo, který provedl disekci i traktografii mozků kočkodanů (také čeleď kočkodanovití, stejně jako makak) a IFOF zde nalezl [10].
Cíl
Cílem této práce je poskytnout obrazový materiál pro vybudování schopnosti představit si průběh IFOF při pohledu na laterální plochu hemisféry. Tento cíl je prakticky zaměřený, neboť pohled na laterální stranu mozku je pohledem při operacích, při kterých je pacient polohován na boku, což je u neurochirurgických operací vcelku častá poloha. Účelem je vštípit si průběh IFOF do paměti tak, aby při pohledu na laterální plochu hemisféry operatér „viděl“ IFOF v kontextu s rozpoznatelnými anatomickými znaky frontálního, temporálního a parietálního laloku, a mohl tak předvídat, kde se IFOF v operačním poli nachází.
Metoda
Ke studiu anatomického průběhu IFOF byla využita jednak anatomická preparace a jednak DTI traktografie.
Anatomická preparace
Mozek zemřelého dárce byl uložen alespoň na 40 dní v 10% formalínu (případná delší doba není na závadu, naopak). Pak byl mozek vyjmut z formalínu a 24 h ponechaný ponořený v pomalu volně přitékající vodě. Následně byl mozek zmrazen při –15 °C a ponechán takto 2 týdny. Poté byl mozek rozmrazen ve vlažné vodě a uložen zpět do formalínu, tentokrát již jen do 2% roztoku (možno nechat pouze v alkoholovém roztoku).
Tento postup přípravy se nazývá metoda dle Klinglera [4]. Vůči obvyklé fixaci formalínem je zde navíc krok zmrazení. Zmrazení vede k rozvolnění svazků vláken bílé hmoty a jejich snazší preparaci.
Preparace se provádí jednak dřevěnou lopatkou zabroušenou do špičky o síle cca 2–4 mm, kterou se stírá šedá kůra mozková až do dosažení bílé hmoty. Při preparaci bílé hmoty je základním nástrojem tupý (mikro) disektor. Hrotem disektoru je nalezena štěpící se vrstva bílé hmoty, kterou je potřeba „rozčesávat“ na jednotlivé vrstvy představující svazky vláken. Víceméně nutnou pomůckou je také preparační mikroskop.
Nejdříve byl pomocí skalpelu odříznut horní temporální gyrus. Tím se odkryla dolní polovina inzulárního laloku. Pak byla dřevěnou lopatkou setřena kůra inzuly v rozsahu viditelné části inzuly. Pod kůrou inzuly se nachází bílá hmota (capsula extrema, která je podkladem gyrů inzuly). Capsula extrema byla odpreparovávána a v místě pod (již odpreparovaným) apexem inzuly se nachází vlákna, patřící IFOF (a také fasciculus uncinatus).
Po nalezení IFOF bylo podél jeho vláken postupováno s preparací distálně, do místa, kde IFOF vbíhá do temporálního laloku těsně pod dolním periinzulárním sulkem. Postupným odpreparováváním dalších kortikálních oblastí (středního temporálního gyru, angulárního gyru, částečně i supramarginálního gyru) a i vláken bílé hmoty (zejména terminálních částí fasciculus longitudinalis superior a fasciculus arcuatus) tak byl ozřejměn další průběh vláken IFOF přes atrium postranní komory a následně do kortikálních oblastí okcipitálního, parietálního a částečně i temporálního laloku (temporobazálně).
Nakonec bylo postupováno s preparací od inzuly proximálním směrem do frontálního laloku. Vlákna IFOF se zde stáčí na vnitřní stranu operkul (mimo pars opercularis), kde byly vypreparovány až do kortikálních zakončení. Pars opercularis je vlákny IFOF podbíhána, byla tedy odpreparována, aby byla tato vlákna jdoucí k dorzálním částem středního a horního frontálního gyru patrná.
DTI rekonstrukce
Difuzí vážené zobrazení MR (diffusion-weighted imaging; DWI) sekvence byly získány v Laboratoři multimodálního a funkčního zobrazování v Central European Institute of Technology v Brně (CEITEC), zpracováno bylo jedno vyšetření dobrovolníka bez patologického nálezu na MR. Vyšetření bylo provedeno na MR Siemens Prisma 3T. Snímáno bylo 92 axiálních skenů při 64 směrech difuzních gradientů (b = 1000 s/ mm2). Parametry obrazu byly následující: TR = 3230 ms, TE = 89,2 ms, sklápěcí úhel = 78°, akviziční matrix 138 × 138, velikost voxelu 1,5 mm × 1,5 mm × 1,5 mm. Tato série 64 směrů gradientů byla vyšetřena dvakrát, vždy s opačným směrem fázového kódování (anterior posterior [AP] a posterior anterior [PA]). Pro každý směr (AP a PA) byla vždy doplněna série s b = 0 s/ mm2.
Získaná data byla předzpracována za použití nástroje FSL [11] – FSL TOPUP pro korekci geometrických zkreslení vzniklých magnetickou susceptibilitou a FSL EDDY pro korekci zkreslení vzniklých vířivými proudy a pohyby hlavy.
Traktografie byla prováděna programem MedInria (Inria, Le Chesnay-Rocquencourt, Francie) [12], v kterém jsou implementovány algoritmy pro analýzu dat DWI. Traktografie je v MedInrii implementována za použití DTI. Nejdříve byla provedena „whole brain tractography“ s parametry „Start FA Treshold“ 300 (odpovídá frakční anizotropii 0,3), „Stop FA Treshold“ 200 (odpovídá frakční anizotropii 0,2), „Smoothness“ 50 (jedná se o specifický parametr algoritmu traktografie použitý v MedInrii, který stanovuje, jak se má algoritmus chovat v místech s nevyhraněnou anizotropií, jak bylo popsáno podrobněji v [13], a „Minimum lenght“ 100 (minimální délka traktu v mm). Poté byla pomocí definice „inclusion“ ROI (region of interest) vybrána pouze vlákna procházející přes tzv. „temporal stem“, tj. místo, kde je temporální lalok fixován k bazi frontálního laloku. Vlákna IFOF jsou zde velmi koncentrovaná, takže je snadné zde celý svazek zachytit (obr. 1G).
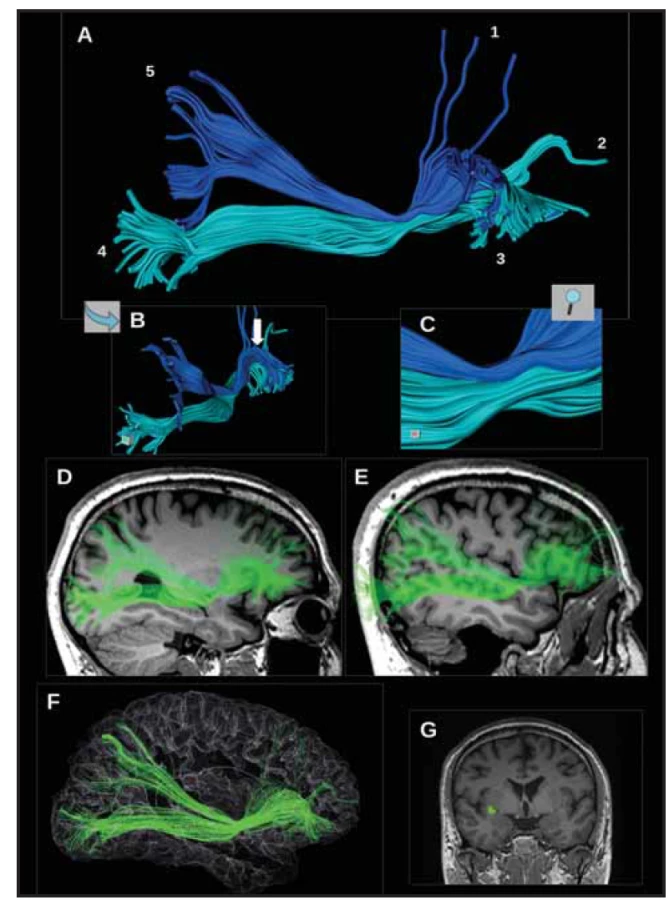
(A) Třírozměrný pohled na pravostranný
IFOF z laterální strany. Tmavě
modře je zobrazena dorzální větev,
světle modře ventrální větev. Kortikální
zakončení odpovídají následujícím
oblastem: 1 – horní a střední frontální
gyrus (DLPFC); 2 – pól frontálního
laloku; 3 – kůra laterálního a zadního
orbitálního gyru a pars orbitalis dolního
frontálního gyru; 4 – pars orbitalis
a pars triangularis; 5 – struktrury okcipitálního
laloku; 6 – horní parietální
lobulus.
(B) Pohled směrem od okcipitálního laloku,
kde jde dobře vidět klenutí IFOF
laterálně směrem k pars triangularis
a pars orbitalis (šipka).
(C) Pohled na kmen IFOF s oběma větvemi.
Traktografií se zřetelně oddělují
obě větve IFOF a to především
dorzálně od kmene IFOF, směrem k okcipitálnímu
a parietálnímu laloku. Ventrálně
od kmene IFOF do sebe kortikální
zakončení obou větví plynule
přecházejí v oblasti dolního frontálního
gyru. Traktografie podhodnocuje
množství kortikálních zakončení
v DLPFC, je to zřejmě důsledek použitého
algoritmu, vycházejícího ze zobrazení
tenzorů difuze.
(D) IFOF vede ve stropu a v laterální
stěně temporálního rohu postranní komory
a přechází na laterální stěnu atria
postranní komory.
(E, F) Vztah IFOF a kortikálních struktur
laterální plochy hemisféry.
(G) Místo, kde prochází kmen IFOF
přes tzv. „temporal stem“, kde je
vhodné začít se segmentací IFOF při
traktografi i.
IFOF – fasciculus fronto-occipitalis
inferior
Fig. 1. Results of tractographic reconstructions of the IFOF.
(A) Three-dimensional view of the right-sided IFOF from the lateral side. The dorsal branch is shown in dark blue, and the ventral
branch in light blue. Cortical terminations correspond to the following areas: 1 – superior and middle frontal gyri (DLPFC); 2 – pole
of the frontal lobe; 3 – cortex of the lateral and posterior orbital gyri and orbital part of the inferior frontal gyrus; 4 – orbital part and
triangular part; 5 – occipital lobe; 6 – superior parietal lobule.
(B) View from the occipital lobe. The arch of the IFOF turning laterally towards the triangular and orbital parts (arrow) can be seen.
(C) View of the IFOF stem with both branches. Tractography clearly distinguishes the two branches of the IFOF, especially dorsally
from the IFOF stem, towards the occipital and parietal lobes. Ventrally from the IFOF stem, the cortical terminations of the two branches
merge smoothly in the area of the inferior frontal gyrus. Tractography underestimates the number of cortical terminations in the
DLPFC. This is probably due to the used algorithm based on diff usion tensor imaging.
(D) The IFOF courses in the roof and in the lateral wall of the temporal horn of the lateral ventricle and passes to the lateral wall of the
atrium of the lateral ventricle.
(E, F) Relationship between the IFOF and cortical structures of the lateral surface of the hemisphere.
(G) The place where the IFOF stem passes through the so-called “temporal stem”. It is a suitable location to start the segmentation of
the IFOF during tractography.
IFOF – inferior fronto-occipital fascicle
Takto vybraný svazek však obsahuje řadu vláken, která k IFOF nepatří, takže tyto byly následně odstraněny (vytvořením příslušných „exclusion“ ROI). Jedná se zejména o vlákna fasciculus uncinatus jdoucí do pólu temporálního laloku, o vlákna jdoucí do horního a středního temporálního gyru (která také pravděpodobně (viz v Diskuzi a obr. 2 A, B) nenáleží k IFOF) a dále pak o vlákna zjevně arteficiální, tj. např. překračující střední čáru.
(B) Vzhledem k velmi pravděpodobnému zapojení těchto vláken do sémantiky a vzhledem k průběhu těchto vláken nacházejících se v bezprostřední
blízkosti IFOF by bylo logické považovat tyto spoje za součást IFOF. Avšak při preparaci popsaným postupem (se započetím preparace
v kmeni IFOF) jsou tato vlákna při preparaci oddělena od IFOF a IFOF pak nemá kortikální zakončení v laterálních oblastech temporálního laloku.
Tato vlákna pak anatomicky přináleží k vláknům capsuly extremy.
(C) Fotografie z původního Curranova popisu IFOF. Je zde zřetelně vidět oddělení IFOF od okolních struktur, tj. od putamen dorzálně a od fasciculus
uncinatus ventrálně. Toto zřetelné oddělení vybízí ke zvolenému postupu, totiž považovat za IFOF vše, co je právě v tomto „kmenu“.
(D) Při traktografi i není nikdy jisté, zda jsou zobrazené struktury skutečné nebo ne, což má ilustrovat tento obrázek. Na obrázku vidíme IFOF
(modře) a fasciculus longitudalis superior (žlutě). Tmavě červenou barvou jsou vyznačena vlákna, která se pravidelně na traktografi i zobrazují
a která jdou při horním okraji IFOF a stáčí se po předním dolním okraji fasciculus longitudinalis superior směrem k dorzálním oblastem temporálního
laloku. Tato vlákna jsou v literatuře označována za artefakt [28], avšak autoři tohoto článku se domnívají, že by možná skutečná být mohla.
IFOF – fasciculus fronto-occipitalis inferior
Fig. 2. (A) Although the IFOF, as it can be dissected and also depicted with using the tractography, corresponds well with the clinically known
sites involved in speech semantics, an important cortical area where speech semantics are often localized is not within the reach of the IFOF.
These are the dorsal areas of the upper and middle temporal gyri, where semantic disorders in awake craniotomies during electrical stimulation
are often found. Tractography can show connections emanating from the posterior parts of the upper and middle temporal gyri going to
the orbital part of the inferior frontal gyrus.
(B) Given that the involvement of these fi bers in the semantics is very likely and given the course of these fi bers, which are located in the immediate
vicinity of the IFOF, it would be consistent to consider these connections as a part of the IFOF. However, when dissection is done as described
(starting with preparation in the IFOF stem), these fi bers are separated from the IFOF during dissection, and the IFOF then has no cortical
termination in the lateral regions of the temporal lobe. These fi bers anatomically belong to the fi bers of the extreme capsule.
(C) Photographs from Curran‘s original IFOF description. The separation of IFOF from the surrounding structures, i.e., dorsally from the putamen
and ventrally from the uncinate fascicle, is clearly visible. This obvious separation supports the described method of dissection, namely to consider
IFOF as everything that is in this „stem“.
(D) This picture is intended to illustrate that in terms of the tractography results, it is never certain whether the depicted tracts are real or not.
In this picture, we can see the IFOF (blue) and the superior longitudinal fascicle (yellow). There are fi bers, regularly depicted on the tractography
image, which run on the upper edge of the IFOF and twist along the anterior lower edge of the superior longitudinal fascicle towards the
dorsal areas of the temporal lobe (dark red). These fi bers are referred to in the literature as an artifact [28], but the authors of this article believe
that maybe they are real.
IFOF – inferior fronto-occipital fascicle
![Obr. 2. (A) Ačkoliv IFOF, tak jak jej lze vypreparovat a také i zobrazit na traktografi i, dobře koresponduje s klinicky známými místy podílejícími
se na sémantice řeči, důležitá kortikální area, kde sémantika řeči bývá často lokalizovaná, v dosahu IFOF není. Jedná se o dorzální oblasti horního
a středního temporálního gyru, kde bývají sémantické poruchy při awake kraniotomiích při elektrické stimulaci nezřídka nalezeny. Traktografií
lze zobrazit spoje vycházející ze zadních částí horního a středního temporálního gyru jdoucí do pars orbitalis dolního frontálního gyru.<br>
(B) Vzhledem k velmi pravděpodobnému zapojení těchto vláken do sémantiky a vzhledem k průběhu těchto vláken nacházejících se v bezprostřední
blízkosti IFOF by bylo logické považovat tyto spoje za součást IFOF. Avšak při preparaci popsaným postupem (se započetím preparace
v kmeni IFOF) jsou tato vlákna při preparaci oddělena od IFOF a IFOF pak nemá kortikální zakončení v laterálních oblastech temporálního laloku.
Tato vlákna pak anatomicky přináleží k vláknům capsuly extremy.<br>
(C) Fotografie z původního Curranova popisu IFOF. Je zde zřetelně vidět oddělení IFOF od okolních struktur, tj. od putamen dorzálně a od fasciculus
uncinatus ventrálně. Toto zřetelné oddělení vybízí ke zvolenému postupu, totiž považovat za IFOF vše, co je právě v tomto „kmenu“.<br>
(D) Při traktografi i není nikdy jisté, zda jsou zobrazené struktury skutečné nebo ne, což má ilustrovat tento obrázek. Na obrázku vidíme IFOF
(modře) a fasciculus longitudalis superior (žlutě). Tmavě červenou barvou jsou vyznačena vlákna, která se pravidelně na traktografi i zobrazují
a která jdou při horním okraji IFOF a stáčí se po předním dolním okraji fasciculus longitudinalis superior směrem k dorzálním oblastem temporálního
laloku. Tato vlákna jsou v literatuře označována za artefakt [28], avšak autoři tohoto článku se domnívají, že by možná skutečná být mohla.<br>
IFOF – fasciculus fronto-occipitalis inferior<br>
Fig. 2. (A) Although the IFOF, as it can be dissected and also depicted with using the tractography, corresponds well with the clinically known
sites involved in speech semantics, an important cortical area where speech semantics are often localized is not within the reach of the IFOF.
These are the dorsal areas of the upper and middle temporal gyri, where semantic disorders in awake craniotomies during electrical stimulation
are often found. Tractography can show connections emanating from the posterior parts of the upper and middle temporal gyri going to
the orbital part of the inferior frontal gyrus.<br>
(B) Given that the involvement of these fi bers in the semantics is very likely and given the course of these fi bers, which are located in the immediate
vicinity of the IFOF, it would be consistent to consider these connections as a part of the IFOF. However, when dissection is done as described
(starting with preparation in the IFOF stem), these fi bers are separated from the IFOF during dissection, and the IFOF then has no cortical
termination in the lateral regions of the temporal lobe. These fi bers anatomically belong to the fi bers of the extreme capsule.<br>
(C) Photographs from Curran‘s original IFOF description. The separation of IFOF from the surrounding structures, i.e., dorsally from the putamen
and ventrally from the uncinate fascicle, is clearly visible. This obvious separation supports the described method of dissection, namely to consider
IFOF as everything that is in this „stem“.<br>
(D) This picture is intended to illustrate that in terms of the tractography results, it is never certain whether the depicted tracts are real or not.
In this picture, we can see the IFOF (blue) and the superior longitudinal fascicle (yellow). There are fi bers, regularly depicted on the tractography
image, which run on the upper edge of the IFOF and twist along the anterior lower edge of the superior longitudinal fascicle towards the
dorsal areas of the temporal lobe (dark red). These fi bers are referred to in the literature as an artifact [28], but the authors of this article believe
that maybe they are real.<br>
IFOF – inferior fronto-occipital fascicle](https://www.csnn.eu/media/cache/resolve/media_object_image_large/media/image_pdf/15cc3dc8b8b231b2a3c96e30d365ed86.png)
Kromě zmíněných programů MedInria a FLS byly ke zpracování výsledků použity programy 3DSlicer [14] a MRIcroGL (Nitrc, University of Massachusetts Medical School in Worcester, MA, USA) [15]. V obr. 3 A, B je využita referenční mapa MNI152 (Montreal Neurological Institute) [16].
(B) Pohled na inzulu po „odpreparování“ opercul. Pod hrotem pars triangularis se nachází apex inzuly (Ap). Kaudálně od apexu je limen
inzulae (L, limen = práh), což je val, který překonává arteria cerebri media cestou do regionu inzuly. Od apexu kraniálně vycházejí krátké
gyry inzuly (SG) a dorzálně od nich je sulcus centralis insulae. Ještě dorzálněji jsou dlouhé gyry inzuly (LG), které se sbíhají k limen. Od
okolních laloků je inzula oddělena horním, dolním a předním periinzulárním sulkem. V kresbě inzuly je červenou barvou znázorněna
projekce putamen.
(C) Ačkoliv je IFOF jednou z nejdůležitějších asociační drah bílé hmoty, je jen jednou z několika popisovaných. Pro orientační představu
o ostatních dráhách jsou na obrázku znázorněny další důležité trakty. Jedná se o corona radiata (tmavě modře, součástí corona radiata
je kortikospinální dráha), fasciculus longitudinalis superior (žlutě), fasciculus arcuatus (světle modře), fasciculus longitudinalis inferior
(fi alově) a fasciculus uncinatus (oranžově).
IFOF – fasciculus fronto-occipitalis inferior
Fig. 3. (A) The most noticeable structure of the lateral surface of the hemisphere is the Sylvian fissure. Above its anterior part lies the
inferior frontal gyrus consisting of parts called the opercular part (Op), triangular part (Tr), and orbital part (Or). During surgery, the easiest
way to recognize these parts is via the triangular part, which has a triangular shape and the branch of the inferior frontal sulcus runs
into it from above. Its tip tends to be somewhat retracted [16], which enlarges the subarachnoid space under the tip. The enlargement is
usually covered with small veins, so a typical „blue spot“ is noticeable here, which is a good reference point. Both the opercular part and
triangular part cover the anterior upper part of the insula. The orbital part is topographically located in front of the insula and continues
into the lateral and posterior orbital gyrus. The opercular part is connected by a typical arch („U“ [16]) to the lower part of the precentral
gyrus. The middle frontal gyrus (MFG) runs parallel to the inferior frontal gyrus. The superior frontal sulcus separates the superior (SFG)
and middle frontal gyri. It is more or less continuous and clearly visible during the surgery. It ends dorsally directly in front of the socalled
Ω region, which is the primary motor area of the hand on the precentral gyrus (PG). On the lateral surface of the hemisphere there
are also gyri of the temporal lobe – superior (STG), middle (MTG), and inferior temporal gyri (ITG). The upper temporal gyrus covers the
lower part of the insula. Slightly behind half of the superior temporal gyrus is the transversely oriented Heschl‘s gyrus (He, primary
auditory area). The postcentral gyrus (POG) lies over the Heschl‘s gyrus [16]. The axis of the Heschl‘s gyrus points to the atrium of the
lateral ventricle [16]. The posterior end of the Sylvian fi ssure turns cranially, and the supramarginal gyrus (SG) is wrapped around this
posterior branch of the Sylvian fi ssure. Dorsally and caudally from the supramarginal gyrus there is the area of the angular gyrus (AG).
The extension of the superior temporal sulcus continues into the area of the angular gyrus. The angular gyrus and the supramarginal
gyrus form the lower parietal lobule. The lower parietal lobule is separated from the upper parietal lobule by the intraparietal sulcus (I).
The boundary between the angular and supramarginal gyri is formed by the intermediate sulcus. The boundary between the occipital
lobe, the parietal lobe, and the temporal lobe is given arbitrarily. The lateral surface of the occipital lobe consists of three gyri (inferior
[IOG], medial [MOG] and superior occipital gyri [SOG]), but their anatomical structure is individually highly variable. DLPFC (dorsolateral
prefrontal cortex) is an important cortical area where, among the other tracts, the IFOF terminates and where during awake
craniotomies many IFOF-related functions can be disturbed by the electrical stimulation.
(B) View of the insula after „dissection“ of the opercula. Underneath the tip of the triangular part there is the apex of the insula (Ap).
Caudally from the apex there is the limen insulae (L, limen = threshold), which is the edge where the middle cerebral artery crosses on
its way to the region of the insula. The short gyri of the insula (SG) run cranially from the apex. Dorsally from the short gyri is the central
sulcus of the insula. More dorsally there are the long gyri of the insula (LG), which converge to the limen. The insula is separated from
the surrounding lobes by an upper, lower, and anterior periinsular sulci. The projection of the putamen is shown in red in the drawing
of the insula.
(C) Although the IFOF is one of the most important white matter association tracts, it is only one of several tracts described. For an
approximate idea of the other bundles, the fi gure shows the other important tracts. These are the corona radiata (dark blue, the part
of the corona radiata is the corticospinal tract), the superior longitudinal fascicle (yellow), the arcuate fascicle (light blue), the inferior
longitudinal fascicle (purple), and the uncinate fascicle (orange).
IFOF – inferior fronto-occipital fascicle
![Obr. 3. (A) Nejvýraznější strukturou laterálního povrchu hemisféry je Sylvická rýha. Nad její přední částí leží dolní frontální gyrus
sestávající se z částí zvaných pars opercularis (Op), pars triangularis (Tr) a pars orbitalis (Or). Peroperačně nejsnáze rozpoznatelná z nich je
pars triangularis, která má trojúhelnikovitý tvar a shora do ní vbíhá větev dolního frontálního sulku. Její hrot bývá poněkud retrahovaný
[16], což rozšiřuje subarachnoidální prostor. Rozšíření bývá překryto drobnými žilami, a tak zde vzniká typická „modrá skrvrna“, která
je dobrým orientačním bodem. Pars opercularis a pars triangularis kryjí přední horní část inzuly. Pars orbitalis se topografi cky nachází
již před inzulou a plynule přechází do laterálního a do zadního orbitálního gyru. Pars opercularis je typickým obloučkem („U“ [16])
propojena s dolní části precentrálního gyru (PG). Paralelně s dolním frontálním gyrem vede střední frontální gyrus (MFG). Horní frontální
sulkus odděluje horní (SFG) a střední frontální gyrus. Bývá víceméně spojitý a při operaci dobře patrný. Dorzálně končí přímo před
tzv. Ω regionem [16], což je primární motorická oblast ruky na precentrálním gyru. Na laterálním povrchu hemisféry se dále nachází
gyry temporálního laloku – horní (STG), střední (MTG) a dolní temporální gyrus (ITG). Horní temporální gyrus kryje dolní část inzuly.
Lehce za polovinou horního temporálního gyru se nachází transverzálně orientovaný Heschlův závit (He, primární sluchová oblast).
Přímo na Heschlův závit nasedá pak postcentrální gyrus (POG [16]). Osa Heschlova závitu směřuje do atria postranní komory [16]. Zadní
konec Sylvické brázdy se zalamuje kraniálně a kolem této zadní větve Sylvické brázdy je obtočen supramarginální gyrus (SG). Dorzálně
a kaudálně od supramarginálního gyru se nachází area angulárního gyru (AG). Do plochy angulárního gyru pokračuje v prodloužení
horní temporální sulkus. Gyrus angularis a gyrus supramarginalis tvoří dolní parietální lobul. Dolní parietální lobul je oddělený od
horního parietálního lobulu intraparietálním sulkem (I). Hranici mezi angulárním a supramarginálním gyrem tvoří intermediární sulkus.
Hranice mezi okcipitálním lalokem, parietálním lalokem a temporálním lalokem je dána arbitrálně. Laterální povrch okcipitálního laloku
se skládá ze tří gyrů (gyrus occipitalis inferior [IOG], medius [MOG], superior [SOG]), jejich anatomická stavba je ale individuálně dosti
proměnlivá. Jako DLPFC (dorso-lateral prefrontal cortex) je označena důležitá kortikální oblast, kde má mimo jiné zakončení IFOF a kde
lze typicky při awake kraniotomiích elektrickou stimulací vyvolat poruchy řady funkcí s IFOF spjatých.<br>
(B) Pohled na inzulu po „odpreparování“ opercul. Pod hrotem pars triangularis se nachází apex inzuly (Ap). Kaudálně od apexu je limen
inzulae (L, limen = práh), což je val, který překonává arteria cerebri media cestou do regionu inzuly. Od apexu kraniálně vycházejí krátké
gyry inzuly (SG) a dorzálně od nich je sulcus centralis insulae. Ještě dorzálněji jsou dlouhé gyry inzuly (LG), které se sbíhají k limen. Od
okolních laloků je inzula oddělena horním, dolním a předním periinzulárním sulkem. V kresbě inzuly je červenou barvou znázorněna
projekce putamen.<br>
(C) Ačkoliv je IFOF jednou z nejdůležitějších asociační drah bílé hmoty, je jen jednou z několika popisovaných. Pro orientační představu
o ostatních dráhách jsou na obrázku znázorněny další důležité trakty. Jedná se o corona radiata (tmavě modře, součástí corona radiata
je kortikospinální dráha), fasciculus longitudinalis superior (žlutě), fasciculus arcuatus (světle modře), fasciculus longitudinalis inferior
(fi alově) a fasciculus uncinatus (oranžově).<br>
IFOF – fasciculus fronto-occipitalis inferior<br>
Fig. 3. (A) The most noticeable structure of the lateral surface of the hemisphere is the Sylvian fissure. Above its anterior part lies the
inferior frontal gyrus consisting of parts called the opercular part (Op), triangular part (Tr), and orbital part (Or). During surgery, the easiest
way to recognize these parts is via the triangular part, which has a triangular shape and the branch of the inferior frontal sulcus runs
into it from above. Its tip tends to be somewhat retracted [16], which enlarges the subarachnoid space under the tip. The enlargement is
usually covered with small veins, so a typical „blue spot“ is noticeable here, which is a good reference point. Both the opercular part and
triangular part cover the anterior upper part of the insula. The orbital part is topographically located in front of the insula and continues
into the lateral and posterior orbital gyrus. The opercular part is connected by a typical arch („U“ [16]) to the lower part of the precentral
gyrus. The middle frontal gyrus (MFG) runs parallel to the inferior frontal gyrus. The superior frontal sulcus separates the superior (SFG)
and middle frontal gyri. It is more or less continuous and clearly visible during the surgery. It ends dorsally directly in front of the socalled
Ω region, which is the primary motor area of the hand on the precentral gyrus (PG). On the lateral surface of the hemisphere there
are also gyri of the temporal lobe – superior (STG), middle (MTG), and inferior temporal gyri (ITG). The upper temporal gyrus covers the
lower part of the insula. Slightly behind half of the superior temporal gyrus is the transversely oriented Heschl‘s gyrus (He, primary
auditory area). The postcentral gyrus (POG) lies over the Heschl‘s gyrus [16]. The axis of the Heschl‘s gyrus points to the atrium of the
lateral ventricle [16]. The posterior end of the Sylvian fi ssure turns cranially, and the supramarginal gyrus (SG) is wrapped around this
posterior branch of the Sylvian fi ssure. Dorsally and caudally from the supramarginal gyrus there is the area of the angular gyrus (AG).
The extension of the superior temporal sulcus continues into the area of the angular gyrus. The angular gyrus and the supramarginal
gyrus form the lower parietal lobule. The lower parietal lobule is separated from the upper parietal lobule by the intraparietal sulcus (I).
The boundary between the angular and supramarginal gyri is formed by the intermediate sulcus. The boundary between the occipital
lobe, the parietal lobe, and the temporal lobe is given arbitrarily. The lateral surface of the occipital lobe consists of three gyri (inferior
[IOG], medial [MOG] and superior occipital gyri [SOG]), but their anatomical structure is individually highly variable. DLPFC (dorsolateral
prefrontal cortex) is an important cortical area where, among the other tracts, the IFOF terminates and where during awake
craniotomies many IFOF-related functions can be disturbed by the electrical stimulation.<br>
(B) View of the insula after „dissection“ of the opercula. Underneath the tip of the triangular part there is the apex of the insula (Ap).
Caudally from the apex there is the limen insulae (L, limen = threshold), which is the edge where the middle cerebral artery crosses on
its way to the region of the insula. The short gyri of the insula (SG) run cranially from the apex. Dorsally from the short gyri is the central
sulcus of the insula. More dorsally there are the long gyri of the insula (LG), which converge to the limen. The insula is separated from
the surrounding lobes by an upper, lower, and anterior periinsular sulci. The projection of the putamen is shown in red in the drawing
of the insula.<br>
(C) Although the IFOF is one of the most important white matter association tracts, it is only one of several tracts described. For an
approximate idea of the other bundles, the fi gure shows the other important tracts. These are the corona radiata (dark blue, the part
of the corona radiata is the corticospinal tract), the superior longitudinal fascicle (yellow), the arcuate fascicle (light blue), the inferior
longitudinal fascicle (purple), and the uncinate fascicle (orange).<br>
IFOF – inferior fronto-occipital fascicle](https://www.csnn.eu/media/cache/resolve/media_object_image_large/media/image_pdf/c471caf7e4e3f4212b15f08431df7be3.png)
Výsledky
Aby bylo možno uvést poznatky z práce do patřičného kontextu, jsou na obr. 3 shrnuty potřebné anatomické základy.
Fotografie IFOF pořízené při preparaci jsou na obr. 4. Na obr. 1 jsou pak traktografické rekonstrukce. Dále uvedené poznatky jsou syntézou nálezů získaných při disekci i při traktografii.
(B, C) Zvětšenina zobrazující průběh dorzální (horní) větve IFOF (po odpreparování pars opercularis) směrem k dorzálním kortikálním
oblastem horního a středního frontálního gyru a na obrázku C pak vyústění těchto vláken na kůře. Jedná se vlastně o vyústění na tzv.
dorzo-laterálním prefrontálním kortexu, kde velmi často při operacích vídáme funkční odpovědi při stimulaci.
(D) Detaily kortikáních zakončení ventrální (dolní) větve IFOF na kůře pars orbitalis a pars triangularis, poté co se stáčí laterálním směrem
po podběhnutí horního periinzuárního sulku. Rovněž v těchto oblastech vyvolává někdy při awake kraniotomiích elektrická stimulace
funkční poruchy.
(E) Detail průběhu IFOF inzulou v místě před putamen (které je dosud kryto bílou hmotou tvořenou claustro-kortikálními vlákny)
(F) IFOF v preparátu na obrázku A má pouze omezené množství kortikálních zakončení v gyrus occipitalis inferior a žádná v gyrus
occipitalis medius. To je docela neobvyklé. Ke správné interpretaci tohoto nálezu je nutné vědět, že kvalita preparátu mozku se
v různých místech vždy liší. Obecně, struktury blíže k povrchu mozku bývají tužší a disekce je snadná. V hlubších částech preparátu jsou
však některé části někdy velmi křehké. Není tedy možné vyloučit, že navzdory veškerému úsilí je toto omezené kortikální zakončení
v dolním okcipitálním gyru a chybějící kortikální zakončení ve středním okcipitálním gyru artefaktem. Na obrázku E ale vidíme fotografi i
jiného preparátu, který obsahuje tato typická kortikální zakončení v dolním a středním okcipitálním gyru.
(G) Fotografi e preparátu téhož mozku jako na obrázku A mozku pořízená před začátkem preparace s barevně zvýrazněným průběhem
IFOF (na levé straně). Je tak patrný průmět IFOF do původních kortikálních oblastí.
IFOF – fasciculus fronto-occipitalis inferior
Fig. 4. (A) Photograph of the brain with the IFOF dissected. The fibers of the IFOF are located between the red and blue marks. The IFOF‘s
fi bers tend to split into two components [17,18] creating (especially towards the occipital and parietal lobes) two branches – the more
ventrally (between the blue and green marks) caudally located branch has its cortical terminations in the inferior and middle occipital
gyri and in the temporobasal areas and the more dorsally and cranially located branch (between the green and red marks) has cortical
terminations in the parietal cortical areas and in the superior occipital gyrus. In the direction toward the frontal lobe, the dorsal (upper)
branch tends to lead to the dorsal cortical areas of the superior and medial frontal gyri, and the ventral (inferior) branch runs more
toward the structures of the inferior frontal gyrus (triangular part, orbital part) and to the pole of the frontal lobe, but the branches
overlap to some extent and are not as distinctly separated as in the dorsal direction.
(B, C) Enlargement showing the course of the dorsal (upper) branch of the IFOF (after dissection of the opercular part) towards the
dorsal cortical areas of the superior and middle frontal gyri and at C the termination of these fi bers is on the cortex. This is actually
the termination on the so-called dorso-lateral prefrontal cortex, where functional disturbances can often be seen during electrical
stimulation during awake craniotomies.
(D) Details of the cortical terminations of the ventral (lower) branch of the IFOF are on the cortex of the orbital part and triangular part,
after the fi bers turn laterally after passing under the superior periinsular sulcus. In these areas electrical stimulation during awake
craniotomies sometimes causes functional disorders as well.
(E) Details of the course of the IFOF in the region of the insula, in front of the putamen (in the picture the putamen is still covered by
white matter formed by the claustro-cortical fi bers)
(F) The IFOF in dissection in figure A has only a limited cortical termination in the inferior occipital gyrus and none in the middle occipital
gyrus. This is quite unusual. To interpret this, it necessary to know that the quality of the specimen always varies in its various locations. In
general, closer to the brain surface, the structures tend to be stiff er and the dissection is easier. In deeper parts of the specimen, however,
some parts are sometimes very fragile. Thus, it is not possible to exclude that, despite all eff orts, this limited cortical termination in the
inferior occipital gyrus and missing terminations in the middle occipital gyrus are artifact. In the picture E, however, we can see the
photograph of another specimen that contains these typical cortical terminations in the inferior and middle occipital gyri.
(G) The photograph of the exactly same brain as in picture A, taken before the dissection started is fused over the photograph of the
specimen with the dissected IFOF image highlighted (on the left). This shows the projection of the IFOF into the original cortical areas.
IFOF – inferior fronto-occipital fascicle
![Obr. 4. (A) Fotografie mozku s vypreparovaným IFOF. Vlákna IFOF se nachází mezi červenými a modrými značkami. Vlákna IFOF mají
tendenci se dále štěpit do dvou složek [17,18], čímž se vytvoří (zejména směrem k okcipitálnímu a parietálnímu laloku) dvě větve –
ventrální (mezi modrými a zelenými značkami), níže uložená větev má své kortikální zakončení v dolním a středním okcipitálním gyru
a v temporobazálních oblastech a dorzální, výše uložená větev (mezi zelenými a červenými značkami) má kortikální zakončení směřující
do parietálních korových oblastí a do horního okcipitálního gyru. Směrem k frontálnímu laloku má dorzální (horní) větev tendenci
vést k dorzálním kortikálním oblastem horního a středního frontálního gyru a větev ventrální (dolní) vede spíše ke strukturám dolního
frontálního gyru (pars triangularis, pars orbitalis) a k pólu frontálního laloku. Větve se zde ale do jisté míry překrývají a nejsou tak ostře
oddělené jako opačným směrem.<br>
(B, C) Zvětšenina zobrazující průběh dorzální (horní) větve IFOF (po odpreparování pars opercularis) směrem k dorzálním kortikálním
oblastem horního a středního frontálního gyru a na obrázku C pak vyústění těchto vláken na kůře. Jedná se vlastně o vyústění na tzv.
dorzo-laterálním prefrontálním kortexu, kde velmi často při operacích vídáme funkční odpovědi při stimulaci.<br>
(D) Detaily kortikáních zakončení ventrální (dolní) větve IFOF na kůře pars orbitalis a pars triangularis, poté co se stáčí laterálním směrem
po podběhnutí horního periinzuárního sulku. Rovněž v těchto oblastech vyvolává někdy při awake kraniotomiích elektrická stimulace
funkční poruchy.<br>
(E) Detail průběhu IFOF inzulou v místě před putamen (které je dosud kryto bílou hmotou tvořenou claustro-kortikálními vlákny)<br>
(F) IFOF v preparátu na obrázku A má pouze omezené množství kortikálních zakončení v gyrus occipitalis inferior a žádná v gyrus
occipitalis medius. To je docela neobvyklé. Ke správné interpretaci tohoto nálezu je nutné vědět, že kvalita preparátu mozku se
v různých místech vždy liší. Obecně, struktury blíže k povrchu mozku bývají tužší a disekce je snadná. V hlubších částech preparátu jsou
však některé části někdy velmi křehké. Není tedy možné vyloučit, že navzdory veškerému úsilí je toto omezené kortikální zakončení
v dolním okcipitálním gyru a chybějící kortikální zakončení ve středním okcipitálním gyru artefaktem. Na obrázku E ale vidíme fotografi i
jiného preparátu, který obsahuje tato typická kortikální zakončení v dolním a středním okcipitálním gyru.<br>
(G) Fotografi e preparátu téhož mozku jako na obrázku A mozku pořízená před začátkem preparace s barevně zvýrazněným průběhem
IFOF (na levé straně). Je tak patrný průmět IFOF do původních kortikálních oblastí.<br>
IFOF – fasciculus fronto-occipitalis inferior<br>
Fig. 4. (A) Photograph of the brain with the IFOF dissected. The fibers of the IFOF are located between the red and blue marks. The IFOF‘s
fi bers tend to split into two components [17,18] creating (especially towards the occipital and parietal lobes) two branches – the more
ventrally (between the blue and green marks) caudally located branch has its cortical terminations in the inferior and middle occipital
gyri and in the temporobasal areas and the more dorsally and cranially located branch (between the green and red marks) has cortical
terminations in the parietal cortical areas and in the superior occipital gyrus. In the direction toward the frontal lobe, the dorsal (upper)
branch tends to lead to the dorsal cortical areas of the superior and medial frontal gyri, and the ventral (inferior) branch runs more
toward the structures of the inferior frontal gyrus (triangular part, orbital part) and to the pole of the frontal lobe, but the branches
overlap to some extent and are not as distinctly separated as in the dorsal direction.<br>
(B, C) Enlargement showing the course of the dorsal (upper) branch of the IFOF (after dissection of the opercular part) towards the
dorsal cortical areas of the superior and middle frontal gyri and at C the termination of these fi bers is on the cortex. This is actually
the termination on the so-called dorso-lateral prefrontal cortex, where functional disturbances can often be seen during electrical
stimulation during awake craniotomies.<br>
(D) Details of the cortical terminations of the ventral (lower) branch of the IFOF are on the cortex of the orbital part and triangular part,
after the fi bers turn laterally after passing under the superior periinsular sulcus. In these areas electrical stimulation during awake
craniotomies sometimes causes functional disorders as well.<br>
(E) Details of the course of the IFOF in the region of the insula, in front of the putamen (in the picture the putamen is still covered by
white matter formed by the claustro-cortical fi bers)<br>
(F) The IFOF in dissection in figure A has only a limited cortical termination in the inferior occipital gyrus and none in the middle occipital
gyrus. This is quite unusual. To interpret this, it necessary to know that the quality of the specimen always varies in its various locations. In
general, closer to the brain surface, the structures tend to be stiff er and the dissection is easier. In deeper parts of the specimen, however,
some parts are sometimes very fragile. Thus, it is not possible to exclude that, despite all eff orts, this limited cortical termination in the
inferior occipital gyrus and missing terminations in the middle occipital gyrus are artifact. In the picture E, however, we can see the
photograph of another specimen that contains these typical cortical terminations in the inferior and middle occipital gyri.<br>
(G) The photograph of the exactly same brain as in picture A, taken before the dissection started is fused over the photograph of the
specimen with the dissected IFOF image highlighted (on the left). This shows the projection of the IFOF into the original cortical areas.<br>
IFOF – inferior fronto-occipital fascicle](https://www.csnn.eu/media/cache/resolve/media_object_image_large/media/image_pdf/3ed707bb590d7bafe00ef3044b6ce832.png)
Nejvhodnějším místem k začátku popisu průběhu IFOF je jeho nejužší místo s nejvíce koncentrovanými vlákny (obr. 3C, 4E). Odtud se vlákna postupně rozbíhají oběma směry ke svým kortikálním zakončením. Toto nejužší místo se nachází v inzule, dorzálně od apexu, na přední ploše putamen a bude dále nazýváno kmenem („stem of the IFOF“ v anglicky psaných pracích). V kmeni má IFOF maximální šířku okolo 7 mm. Topograficky se kmen IFOF nachází pod obloučkem (pod „U“) pars opercularis, tj. pod zcela dolní část precentrálního gyru (pozorovatelné na obr. 1E). Směrem dorzálním se IFOF postupně rozšiřuje a zplošťuje. Prochází přitom podél dolního okraje putamen a podbíhá dolní periinzulární sulkus. V místě průchodu se stává součástí stropu temporálního rohu postranní komory, postupně přechází na laterální stěnu temporálního rohu a dorzálněji se plynule stává součástí laterální stěny atria postranní komory (obr. 1D). Temporální roh postranní komory je víceméně horizontální, stejně tak IFOF probíhá v jeho blízkosti horizontálně. Topograficky se horní okraj IFOF víceméně kryje s horním okrajem horního temporálního gyru (obr. 1E) a kraniálně tedy IFOF nedosahuje výšky Sylvické rýhy. Jelikož se ale IFOF plošně vějířovitě rozšiřuje, tak v zadní části středního temporálního gyru již dosahuje přibližně do jeho poloviny.
Přibližně v úrovni konce horizontálního úseku Sylvické brázdy (v místě, kde se zvedá zadní raménko Sylvické fisury) se kraniálně stáčí i IFOF (obr. 4E, 4G) a hluboko pod kůrou supramarginálního gyru a pod dnem intraparietálního sulku (dobře viditelné na obr. 4A) míří do horního parietálního lobulu ke kortikálním zakončením v oblasti lobulus parietalis superior a gyrus occipitalis superior (obr. 1E, 4A, 4G).
Zmíněné kraniální stočení se ale týká horního okraje svazku vláken IFOF. Dolní okraj pokračuje dorzálně v úrovni středního temporálního gyru a jde ke kortikálním zakončením v laterálním povrchu okcipitálního laloku (v oblasti gyrus occipitalis inferior a medius; obr. 1D, 4E), případně také k bazálnímu povrchu hemisféry, k hranici mezi temporálním a okcipitálním lalokem (obr. 1G).
Směrem frontálním od kmene se IFOF taktéž rozšiřuje. Vlákna opouštějí oblast inzuly a podbíhají horní periinzulární sulkus. Dorzální porce vláken se zde klade pod vlákna fasciculus longitudinalis superior a po ohybu (takovém, že stále míří vzhůru, ale šikmo, mediálně) probíhají ke kortikálním zakončením na středním a horním frontálním gyru v jejich dorzální oblastech (obr. 1E, 1F, 4B, 4C). Topograficky takto zasahuje IFOF do úrovně před precentrální gyrus. Ventrální porce vláken se po překonání horního periinzulárního sulku stáčí laterálně a po obloukovitém průběhu ústí na pars triangularis a pars orbitalis dolního frontálního gyru (obr. 4D). Ale ne všechna ventrální vlákna IFOF se stáčejí laterálně do pars triagularis a pars orbitalis. Některá vlákna se laterálně nestočí a procházejí po bazi frontálního laloku až do oblasti pólu frontálního laloku (obr. 1F).
Ve svém úhrnu tak frontálně vlákna IFOF končí na horním, středním a dolním frontálním gyru, a to od pólu až do dorzálních částí, avšak s výjimkou pars opercularis, kam vlákna nevedou. V dorzálních oblastech pak vlákna ústí v horním, středním a dolním okcipitálním gyru, v temporobazálních oblastech v zadní části temporálního laloku a v horním parietálním lobulu.
Diskuze
Anatomie je vědou, u níž důležitou roli hrají definice. Definovat, co je IFOF, ale není snadné.
Nejjednodušeji lze IFOF popsat jako asociační vlákna spojující okcipitální a frontální lalok. Podobně popsal IFOF Curran, který mimo jiné uvádí: „Často jsem nacházel dlouhý svazek sdružujících se vláken spojujících týlní lalok s čelním a procházející po zevní straně a velmi blízko k bazi lentikulárního jádra, v bezprostředním kontaktu s vlákny přední komisury, když vstupují do spánkového laloku. Zpočátku jsem nebyl nakloněn považovat to za spojení týlního laloku s čelním a myslel jsem si, že to pravděpodobně patří k dolním vodorovným vláknům zevní kapsuly. Následné zkoumání to ale vyvrátilo a ukázalo skutečnou podstatu svazku jako souvislých vláken, nepřerušovaných žádnými jádry v po celé délce jeho průběhu.“ [6].
Avšak o kousek dále Curran uvádí: „Přestože většina jeho vláken směřuje přímo do týlního laloku a lze je tam rozpoznat jako víceméně samostatný svazek, některá vlákna se ještě dříve oddělí a končí v kůře zadní části spánkového laloku a některá také v zadní části v temenního laloku.“ [6]. Již Curran tedy rozpoznal, že IFOF obsahuje také vlákna jdoucí do parietálního laloku a temporálního laloku. Není ovšem možné jednoduše říct, že IFOF je tvořen všemi vlákny jdoucími z okcipitálního, temporálního a parietálního laloku do frontálního laloku – tak by totiž zahrnoval i vlákna, která jsou tradičně připisována jiným svazkům.
Jako nejvhodnější k definici IFOF se jeví využít faktu, že v oblasti před putamen lze kmen IFOF velmi snadno oddělit od okolních vláken a struktur (od fasciculus uncinatus, od putamen atd.). Toto nejužší místo je pak považováno za základ IFOF, a tím jsou i jednoznačně dána vlákna, která IFOF patří a která lze sledovat do kortikálních oblastí. Když se opět začteme do Currana, najdeme zmínku, že i on tuto „ochotu“ IFOF oddělit se od okolních struktur patrně pozoroval: „Zdůraznil bych skutečnost, že když obtáčí spodní a vnější stranu lentikulárního jádra a zevní kapsulu, vynikne nápadnou odlišností a dá se okamžitě rozpoznat jako samostatný svazek, izolovaný od okolních struktur přímostí a kompaktností svých vláken.” [6]. Oddělenost IFOF od ostatních struktur zobrazenou Curranem lze pozorovat na obr. 2C.
Samotná vlákna IFOF mají tendenci se dále štěpit do dvou složek, čímž se vytvoří dvě větve, dorzální a ventrální (označení vychází z pořadí uložení větví ve kmeni, dorzální větev je současně kraniálněji uložená), podrobněji viz obr. 4. Toto dělení velmi dobře odpovídá i nálezům na traktografii (obr. 1) a bylo již dříve popisováno [17,18].
Ve svém průběhu přes strop temporálního rohu postranní komory a laterální stěnou atria se IFOF stává součástí tzv. stratum sagittale. Tento pojem zavedl Klingler [4] a jedná se o vrstvu vláken bílé hmoty zahrnující IFOF, vlákna přední komisury a vlákna optické radiace. Nejlaterálněji je uložen IFOF, pod ním jsou vlákna přední komisury a ještě mediálněji vlákna optické radiace. Vlákna optické radiace jsou zdaleka nejpočetnější skupinou ze všech vláken obsažených ve stratum sagittale a v porovnání s optickou radiaci je IFOF pouze gracilní strukturou.
Zavedení jednoho pojmu (tj. stratum sagittale) pro několik skupin vláken odráží fakt, že vlákna jednotlivých systémů jsou zde do značné míry propletená. Je možné separovat např. vlákna IFOF od vláken přední komisury a od vláken optické radiace a vlákna optické radiace od vláken přední komisury, ale je to vždy za cenu poničení části těchto struktur.
Přední komisura je vývojově starou strukturou, která dnes u člověka pravděpodobně nemá funkční význam. Přesto je ale množství vláken, které lze vypreparovat ve stratum sagittale, překvapivě velké. Vlákna lze snadno vystopovat, jak se stávají kompaktním svazkem a mizí v Gratioletově kanálu (Louis Pierre Gratiolet, 1815–1865, francouzský anatom, který jako první postřehl opakující se vzorec sulků a gyrů povrchu mozku a popsal je) pod globus pallidus.
Z praktického (neurochirurgického) hlediska je důležité vědět, že vlákna optické radiace vybíhají z oblasti corpus geniculatum laterale thalamu do stropu temporálního rohu a do laterální stěny postranní komory ventrálně (vpřed), aby se ve svém průběhu vzápětí stočila dorzálně k okcipitálnímu kortexu [19]. Toto stočení je nazýváno Mayerovou kličkou a překrývá temporální roh postranní komory až téměř do jejího pólu. To znamená, že otevření jakékoli části temporálního rohu postranní komory bude mít za následek určité poškození optické radiace (což ale nemusí ihned znamenat klinicky významný výpadek zorného pole).
Přestože byl IFOF již dlouhou dobu znám, teprve v roce 2005 se v literatuře [20] objevila první pozorování týkající se jeho funkce. IFOF je pravděpodobně klíčový pro správnou činnost sémantického systému mozku. Sémantický systém je jedním ze základních prvků kognitivních funkcí. Sestává ze sémantické paměti, ve které jsou informace o vlastnostech objektů vnějšího světa, a z řídicích procesů, které umožňují použití těchto informací k vyvození úsudku [21–23].
Jednou z důležitých vlastností sémantického systému je, že dokáže vytvářet koncepty. Koncept představuje zobecnění objektů, jehož je dosaženo vypuštěním nedůležitých vlastností a ponecháním vlastností charakteristických. Např. nikde na světě neexistují dva přesně stejné stromy, ale přesto má pojem (koncept) „strom“ natolik charakteristické vlastnosti, že je snadné si pod ním představit konkrétní objekt. Pojmenováním konceptů vznikají pojmy a pojmy jsou zase základem pro existenci řeči.
Sémantický systém tak lze rozčlenit na verbální a neverbální. Neverbální zahrnuje utříděnou databázi konceptů a jejich vlastností a verbální sémantický systém pak pro koncept, který je připraven v mysli, dokáže z mentálního slovníku vybrat správný pojem.
Porušení verbálního sémantického systému se typicky projeví sémantickou parafázií – při pojmenování se použije nesprávný (ale jinak smysluplný) pojem, ale objekt s pojmem nějakým způsobem významově souvisí, jako by se v paměti sáhlo těsně vedle. Např. místo hrušky – jablko (blízké druhy ovoce), místo psa – žralok (oba koušou) atd. Poruchy verbálního sémantického systému vznikají při porušení IFOF v dominantní (zpravidla levé) hemisféře. Poruchy neverbální sémantiky vznikají rovněž při porušení IFOF v dominantní hemisféře, ale na rozdíl od verbálních mohou vzniknout trvalé neverbální poruchy i po porušení IFOF v nedominantní hemisféře. Porucha neverbálního sémantického systému se projeví zhoršením úsudku a porozumění okolnímu světu. Ne všichni pacienti s přerušeným IFOF v nedominantní hemisféře trpí poruchou neverbální sémantické kognice, ale z těch pacientů, kteří poruchou trpí, není cca 30 % schopno vrátit se ke své předchozí profesi [24].
Stojí za uvedení, že zatímco dorzální větev IFOF má význam v úkolech vyžadujících porozumění významu (tedy neverbální sémantika), ventrální větev se podílí na automatickém („bezmyšlenkovitém“) pojmenování [25].
Další z funkcí, na nichž se IFOF podílí a která je tak běžná a bezděčná, že si její přítomnost vůbec neuvědomujeme, je schopnost rozpoznat duševní stav druhých lidí, tj. jejich myšlenky, pocity, přesvědčení, záměry [26]. V zahraniční literatuře je pro tuto schopnost používán termín „mentalizing“ anebo také „theory of mind“. V češtině se lze setkat s termínem mentalizace. Schopnost mentalizace má blízko k empatii, ale není to totéž – mentalizace znamená spíše pochopení „rozumem“ než (empatické) vcítění se. Schopnost mentalizace je velmi důležitá pro jakoukoli sociální interakci a pro odpovídající jednání s druhými lidmi. Mentalizace je proces komplexní a nejen IFOF se na správné funkci mentalizace podílí. IFOF má při mentalizaci význam při zrakovém rozeznávání emocí ve tvářích jiných osob [27].
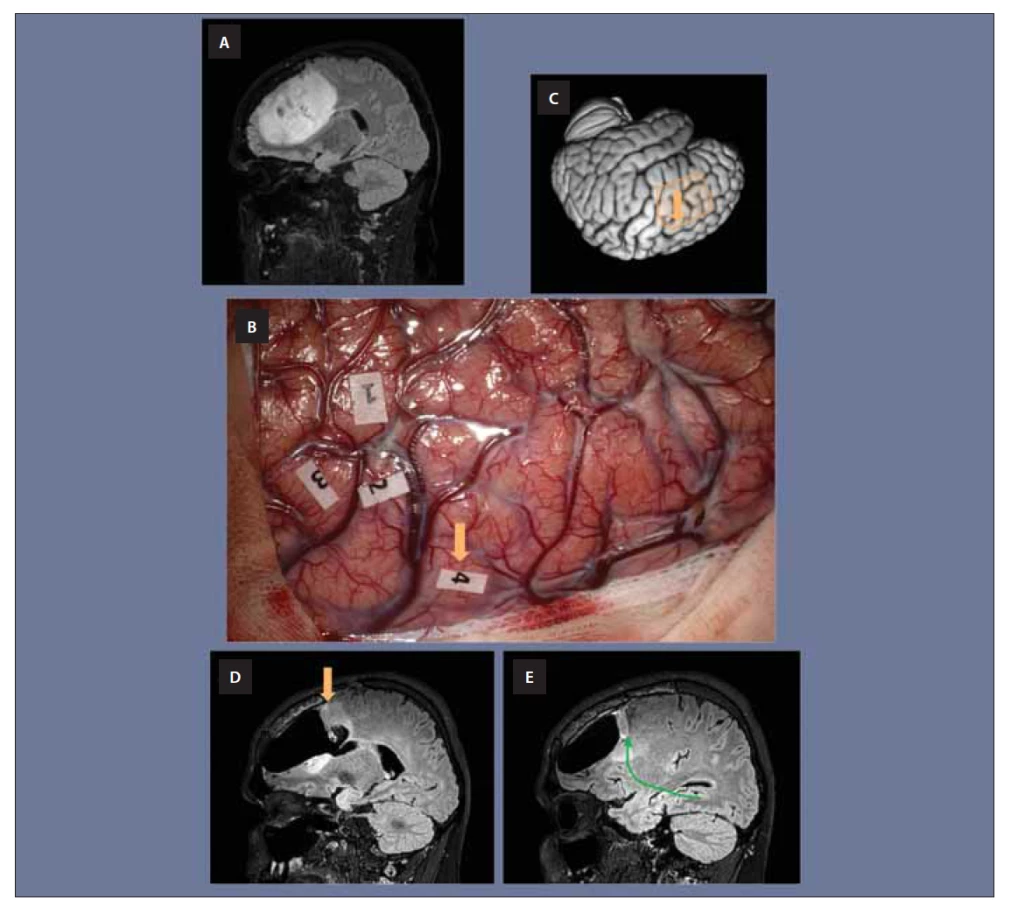
Znalosti funkcí IFOF jsou potřebné při operacích, nejčastěji při operacích gliálních nádorů, a to zejména u awake operací. U awake operací jsou zmiňované funkce testovány. Elektrickou stimulací lze najít funkčně aktivní kortikální oblasti v místě vyústění IFOF (nebo samozřejmě obecně i jiných svazků vláken) anebo v bílé hmotě, zejména v místech, kde jsou vlákna koncentrována do kompaktnějšího svazku. Jeden z příkladů, se kterým se lze v praxi při operaci setkat je popsán na obr. 5.
(B) Snímek z operace této pacientky prováděné v awake kraniotomii. V místech označených čísly 1, 2 a 3 byla vystimulována motorika
obličeje a nacházejí se na precentrálním gyru. V místě s číslem 4 byly elektrickou stimulací vyvolány sémantické parafázie.
(C) Lokalizace operačního pole je schematicky znázorněna na třírozměrném modelu mozku, oranžová šipka odpovídá místu stimulace
(tedy místu pod číslem 4) a nachází se v dorzální části středního temporálního gyru (odpovídá dorzolaterálnímu prefrontálnímu kortexu,
kde vyúsťuje IFOF).
(D) Při operaci zde muselo být ponecháno reziduum tumoru, jež je patrné na pooperační MR.
(E) Zelenou šipkou pak vidíme znázorněný přibližný průběh IFOFu směřující k tomuto místu.
IFOF – fasciculus fronto-occipitalis inferior
Obr. 5. (A) Snímek MR pacientky s rozsáhlým low grade gliomem v levém frontálním laloku.
(B) Snímek z operace této pacientky prováděné v awake kraniotomii. V místech označených čísly 1, 2 a 3 byla vystimulována motorika
obličeje a nacházejí se na precentrálním gyru. V místě s číslem 4 byly elektrickou stimulací vyvolány sémantické parafázie.
(C) Lokalizace operačního pole je schematicky znázorněna na třírozměrném modelu mozku, oranžová šipka odpovídá místu stimulace
(tedy místu pod číslem 4) a nachází se v dorzální části středního temporálního gyru (odpovídá dorzolaterálnímu prefrontálnímu kortexu,
kde vyúsťuje IFOF).
(D) Při operaci zde muselo být ponecháno reziduum tumoru, jež je patrné na pooperační MR.
(E) Zelenou šipkou pak vidíme znázorněný přibližný průběh IFOFu směřující k tomuto místu.
IFOF – fasciculus fronto-occipitalis inferior
Traktografie a disekce jako metody pro studium vláken bílé hmoty mají své přednosti i slabiny. Pokud je při disekci vypreparován nějaký trakt, tak je jisté, že tento nález odpovídá realitě. Vzhledem ke komplexnosti nervových drah a jejich třírozměrnému uspořádání budou ostatní dráhy bohužel nevyhnutelně zničeny, pokud je záměrem vypreparovat jednu určitou. Pokud navíc dojde k chybnému kroku, nelze jej vzít zpět. Traktografie naproti tomu umožňuje zobrazit svazky v jejich třírozměrných vztazích a souvislostech a každou chybu v „preparaci“ lze snadno napravit. Jenže pokud traktografií najdeme nějaká neobvyklá (nečekaná) vlákna, nikdy nevíme, zda jsme objevili něco, co možná ještě nikdo před námi anebo zda se jedná o jeden z bezpočtu artefaktů, kterými je traktografie zatížena (obr. 2D) [28].
Ačkoli se doposud mohlo zdát, že při definici IFOF na základě jeho kmene do sebe vše krásně zapadá, není to tak úplně pravda. Traktografií např. můžeme zobrazit spoje mezi zadní částí středního a horního temporálního gyru a pars orbitalis dolního frontálního gyru. Tato vlákna jsou skutečná, lze je najít i při preparaci. Pokud se řídíme uvedenou definicí IFOF pomocí jeho kmene, pak tato vlákna nepřísluší k IFOF, jelikož se při preparaci oddělí. Víme ale, že se při elektrické stimulaci při awake operacích běžně vyvolá funkční odpověď (anomie, sématická parafázie) v zadní části temporálního laloku (je to koneckonců Wernickeova oblast). Je tak nanejvýš pravděpodobné, že právě tato vlákna mají ve verbální sémantice význam. Zdálo by se tudíž logické (a „elegantní“) z funkčního hlediska, pokud by tato vlákna k IFOF příslušela, avšak morfologicky patří tato vlákna k vláknům capsuly extremy.
Je nutné poznamenat, že fakt, že se v určité oblasti nachází kortikální zakončení vláken IFOF, neznamená automaticky, že by tato oblast musela být klinicky významná a tudíž operačně nedotknutelná. Ale v hloubce bílé hmoty, kde jsou vlákna koncentrovaná do úzkého svazku, je po jejich přerušení funkční deficit vysoce pravděpodobný. Žádná z uvedených funkcí IFOF není testovatelná v celkové anestezii, funkčnímu deficitu se tak lze vyhnout pouze při awake kraniotomii.
V česky psané literatuře se metodou dle Klinglera zabývali autoři Bartoš et al [29] a Hejčl et al [30]. Tyto práce jsou zaměřeny na komplexní a přehledný popis nejdůležitějších drah mediálního i laterálního aspektu hemisféry, tak jak se postupně objevují při disekci. IFOF je v první z nich také zmíněn.
Zvolený postup ostrého odstraňování (odříznutí skalpelem) částí mozku (v této práci zejména části temporálního laloku, ale i jiných okrsků) má výhodu v zachování co nejvíce původních kortikálních struktur a následné dobré orientaci na preparátu. Na druhou stranu je ale preparující tímto postupem ochuzen o možnost vidět další dráhy bílé hmoty, zejména (pokud jde o temporální lalok) fasciculus longitudinalis superior. I tupým nástrojem je možné odpreparovat nejdříve kortex, pak pod kůrou ležící krátká asociační vlákna spojující přilehlé gyry a tak odhalit postupně dlouhé asociační dráhy při postupu latero-mediálně. Lze tak získat lepší představu o vzájemném uspořádání dlouhých asociačních drah. Tento postup je podrobně popsán v nedávno vydané publikaci [31].
A jako drobná poznámka na konec, ačkoli může v první chvíli vyznít podivně: preparáty mozku mají pozoruhodnou estetickou hodnotu, jak lze vidět např. na pracích Richarda G. Párragy [16,19].
Závěr
I v době běžné dostupnosti neuronavigace je stále stěžejní co možná nejlepší znalost anatomie a také funkcí s anatomií spjatých. IFOF sbírá vlákna z rozsáhlých oblastí okcipitálního, parietálního a temporálního laloku a všechna tato vlákna odvádí do laloku frontálního. Není proto překvapující, že se funkčně podílí na celé řadě dějů a že jeho porušení může mít významné funkční dopady. Případné nezamýšlené přetnutí několika milimetrů tkáně navíc může někdy změnit život pacientů k horšímu.
V závěrech článků zpravidla bývá psáno, že další zkoumání je pro lepší objasnění problematiky nutné. Ani tento článek nebude výjimkou. Avšak – inspirováni závěrem v komentáři Marca Cataniho [9] – vypůjčíme si první slova z předmluvy Déjérinovy „Anatomie des centres nerveux“ (1895): „sledování fasciklů, jejich původu, trajektorie a zakončení, to je cíl toho, jenž po Vicq d’Azyrovi (Félix Vicg d’Azyr, 1748–1794, objevitel mamillo-thalamického traktu, osobní lékař poslední francouzské královny Marie Antoinetty) studuje tuto disciplínu anatomie … navzdory zlepšením v technikách, jež jsou nám nyní k dispozici … je zde stále mnoho zastřených bodů k objasnění“.
Grantová podpora
Podpořeno MZ ČR – RVO (FNBr, 65269705)
Etické aspekty
Studie byla provedena ve shodě s Helsinskou deklarací z roku 1975 (a jejími revizemi z let 2004 a 2008).
Konflikt zájmů
Autoři deklarují, že v souvislosti s předmětem studie nemají žádný konflikt zájmů.
Redakční rada potvrzuje, že rukopis práce splnil ICMJE kritéria pro publikace zasílané do biomedicínských časopisů.
The Editorial Board declares that the manuscript met the ICMJE “uniform requirements” for biomedical papers.
MUDr. Ing. Eduard Neuman, Ph.D.
Neurochirurgická klinika FN Brno
Jihlavská 340/20
625 00 Brno
e-mail: neuman.eduard@fnbrno.cz
Přijato k recenzi: 26. 9. 2020
Přijato do tisku: 11. 3. 2021
Sources
1. Sporns O, Tononi G, Kötter R. The human connectome: a structural description of the human brain. PLoS Comput Biol 2005; 1(4): e42. doi: 10.1371/ journal.pcbi.0010042.
2. Duffau H. Brain plasticity: from pathophysiological mechanisms to therapeutic applications. J Clin Neurosci 2006; 13(9): 885–897. doi: 10.1016/ j.jocn.2005.11.045.
3. Ius T, Angelini E, Thiebaut de Schotten M et al. Evidence for potentials and limitations of brain plasticity using an atlas of functional resectability of WHO grade II gliomas: towards a „minimal common brain“. Neuroimage 2011; 56(3): 992–1000. doi: 10.1016/ j.neuroimage.2011.03.022.
4. Ludwig E, Klingler J. Atlas cerebri humani. Basel, N.Y.: Karger 1956.
5. Forkel SJ, Thiebaut de Schotten M, Kawadler JM et al. The anatomy of fronto-occipital connections from early blunt dissections to contemporary tractography. Cortex 2014; 56: 73–84. doi: 10.1016/ j.cortex.2012.09.005.
6. Curran E. A new association fibre tract in the cerebrum. With remarks of the fibre tract dissection method of studying the brain. J Compar Neurol Psychology 1909; 19(6): 1e18.
7. Catani M, Howard RJ, Pajevic S et al. Virtual in vivo interactive dissection of white matter fasciculi in the human brain. Neuroimage 2002; 17(1): 77–94. doi: 10.1006/ nimg.2002.1136.
8. Schmahmann JD, Pandya DN, Wang R et al. Association fibre pathways of the brain: parallel observations from diffusion spectrum imaging and autoradiography. Brain 2007; 130(Pt 3): 630–653. doi: 10.1093/ brain/ awl359.
9. Catani M. From hodology to function. Brain 2007; 130(Pt 3): 602–605. doi: 10.1093/ brain/ awm008.
10. Sarubbo S, Petit L, De Benedictis A et al. Uncovering the inferior fronto-occipital fascicle and its topological organization in non-human primates: the missing connection for language evolution. Brain Struct Funct 2019; 224(4): 1553–1567. doi: 10.1007/ s00429-019-01856-2.
11. FLS. [online]. Available from URL: https:/ / fsl.fmrib.ox.ac.uk/ fsl/ fslwiki/ .
12. medInria. [online]. Available from URL: https:/ / med.inria.fr.
13. Weinstein D, Kindlmann G, Lundberg E. Tensorlines: advection-diffusion based propagation through diffusion tensor fields. San Francisco, CA, USA: Institute of Electrical and Electronics Engineers 1999: 249–530.
14. 3D Slicer. [online]. Available from URL: https:/ / www.slicer.org/ .
15. NITRC. [online]. Available from URL: https:/ / www.nitrc.org/ .
16. Ribas, G. The cerebral architecture. In: Ribas, G. Applied cranial-cerebral anatomy: brain architecture and anatomically oriented microneurosurgery. Cambridge: Cambridge University Press 2018: 15–61.
17. Martino J, Brogna C, Robles SG et al. Anatomic dissection of the inferior fronto-occipital fasciculus revisited in the lights of brain stimulation data. Cortex 2010; 46(5): 691–699. doi: 10.1016/ j.cortex.2009.07.015.
18. Sarubbo S, De Benedictis A, Maldonado IL et al. Frontal terminations for the inferior fronto-occipital fascicle: anatomical dissection, DTI study and functional considerations on a multi-component bundle. Brain Struct Funct 2013; 218(1): 21–37. doi: 10.1007/ s00429 011 0372-3.
19. Párraga RG, Ribas GC, Welling LC et al. Microsurgical anatomy of the optic radiation and related fibers in 3-dimensional images. Neurosurgery 2012; 71 (1 Suppl Operative): 160–172. doi: 10.1227/ NEU.0b013e3182556fde.
20. Duffau H, Gatignol P, Mandonnet E et al. New insights into the anatomo-functional connectivity of the semantic system: a study using cortico-subcortical electrostimulations. Brain 2005; 128(Pt 4): 797–810. doi:10.1093/ brain/ awh423.
21. Lambon Ralph MA. Neurocognitive insights on conceptual knowledge and its breakdown. Philos Trans R Soc Lond B Biol Sci 2013; 369(1634): 20120392. doi: 10.1098/ rstb.2012.0392.
22. Moritz-Gasser S, Herbet G, Duffau H. Mapping the connectivity underlying multimodal (verbal and non-verbal) semantic processing: a brain electrostimulation study. Neuropsychologia 2013; 51(10): 1814–1822. doi: 10.1016/ j.neuropsychologia.2013.06.007.
23. Herbet G, Moritz-Gasser S, Duffau H. Direct evidence for the contributive role of the right inferior fronto-occipital fasciculus in non-verbal semantic cognition. Brain Struct Funct 2017; 222(4): 1597–1610. doi: 10.1007/ s00429-016-1294-x.
24. Duffau H. Miami global brain tumor symposium – Dr. Hugues Duffau. [online]. Available from URL: https:/ / www.youtube.com/ watch?v=Lool6cpkCkc.
25. Rollans C, Cummine J. One tract, two tract, old tract, new tract: a pilot study of the structural and functional differentiation of the inferior fronto-occipital fasciculus. J Neurolinguist 2018; 46: 122–137. doi: 10.1016/ j.jneuroling.2017.12.009.
26. Yordanova YN, Duffau H, Herbet G. Neural pathways subserving face-based mentalizing. Brain Struct Funct 2017; 222(7): 3087–3105. doi: 10.1007/ s00429-017-1388-0.
27. Herbet G, Duffau H. Revisiting the functional anatomy of the human brain: toward a meta-networking theory of cerebral functions. Physiol Rev 2020; 100(3): 1181–1228. doi: 10.1152/ physrev.00033.2019.
28. Fernández-Miranda JC, Rhoton AL Jr, Alvarez-Linera J et al. Three-dimensional microsurgical and tractographic anatomy of the white matter of the human brain. Neurosurgery 2008; 62 (6 Suppl 3): 989–1028. doi: 10.1227/ 01.neu.0000333767.05328.49.
29. Bartoš R, Hejčl A, Zolal A et al. Laboratorní disekce drah laterálního aspektu mozkové hemisféry. Cesk Slov Neurol N 2012; 75/ 108(1): 30–37.
30. Hejčl A, Bartoš R, Zolal A et al. Laboratorní preparace drah z mediálního pohledu na mozkovou hemisféru. Cesk Slov Neurol N 2012; 75/ 108(6): 707–713.
31. Bartoš R, Concepción S. IJ, Němcová V et al. Průvodce mozkem. Neuroanatomie. Praha: Maxdorf 2020.
Labels
Paediatric neurology Neurosurgery NeurologyArticle was published in
Czech and Slovak Neurology and Neurosurgery

2021 Issue 2
Most read in this issue
- Mortonova neuralgie, metatarzalgie
- Nemoc moyamoya
- Správná a chybná pojmenování obrázků pro náročnější test písemného Pojmenování obrázků a jejich vybavení (dveřní POBAV)
- Etiopatogeneze a diagnostika progresivní multifokální leukoencefalopatie u pacientů léčených natalizumabem