
Neurální komponenty motivačních a rozhodovacích procesů v lidském mozku
Neural Components of Motivational and Decision-Making Processes in the Human Brain
Motivational and decision-making processes are important tools in both making daily decisions and overall survival of the physical subject. In the last decade many studies have investigated how these processes function in the human brain. Functional magnetic resonance imaging (fMRI) has played a large part in research into the relationships between motivation and decision-making on the one hand and cerebral activity on the other. Current results suggest that certain areas of the brain are functionally associated with motivational and decision-making processes. This article provides an overview of the most significant recent studies into seeking to identify the structures of the human brain involved in motivational and decision-making processes. In the past, much attention was paid to “gain motivation” (for example, towards financial reward), but current science also focuses on moral motivation, the study of which is addressed in the following part of the article. Some diseases associated with motivational and decision-making disorders are shown in the last part of the article.
Key words:
motivation – decision-making processes – functional magnetic resonance imaging
Autoři:
J. Zelinková 1,2; T. Urbánek 3; R. Mareček 1,2; M. Brázdil 1,2
Působiště autorů:
Výzkumná skupina pro behaviorální a sociální neurovědy, Středoevropský technologický institut (CEITEC), MU, Brno
1; I. neurologická klinika LF MU a FN u sv. Anny v Brně
2; Psychologický ústav Akademie věd ČR
3
Vyšlo v časopise:
Cesk Slov Neurol N 2011; 74/107(4): 419-427
Kategorie:
Přehledný referát
Souhrn
Motivační a rozhodovací procesy představují důležité nástroje jak pro zvládání každodenního života, tak pro přežití člověka obecně. V poslední dekádě byla realizována řada studií, které si kladly za cíl zjistit, jak v lidském mozku tyto procesy fungují. Pro studium vztahů mezi motivací a rozhodováním na jedné a cerebrální aktivitou na druhé straně je používána především funkční magnetická rezonance (fMR). Ze současných poznatků vyplývá, že určité konkrétní oblasti mozku jsou funkčně asociovány s motivačními a rozhodovacími procesy. Předkládaný text poskytuje přehled nejvýznamnějších studií z poslední doby, které si kladly za cíl identifikovat struktury lidského mozku přímo zapojené do motivačních a rozhodovacích procesů. V minulosti byla nejčastěji zkoumána motivace ziskem, například získanou finanční odměnou, ale současná neurověda se zaměřuje také na motivaci morální, jejíž studium je popsáno v další části článku. V poslední části příspěvku jsou pak uvedena některá onemocnění související s poruchami motivačních a rozhodovacích procesů.
Klíčová slova:
motivace – rozhodovací procesy – funkční magnetická rezonance
Úvod
Schopnost rozhodovat se optimálně vzhledem k určité situaci a daným možnostem byla vždy velmi důležitá pro přežití. Rozhodovací procesy souvisí úzce s procesy motivace, které aktivují naše chování a dávají jim účel a směr. Jsou nezbytné pro adaptaci a pro udržování kvality života [1]. Existuje mnoho psychologických teorií motivace a teorií rozhodování [2,3], které ale nejsou tématem tohoto článku. Ten je věnován neurálním projevům, jež je možné prokázat v souvislosti s působením motivačních a rozhodovacích procesů.
Namístě je aspoň stručná psychologická vsuvka. Pojmem motiv se označuje cokoli, co aktivuje a zaměřuje chování lidí. Dále se používají např. pojmy potřeba (buď nedostatek nebo přebytek něčeho), pud (který silně aktivuje chování) a incentiva (pobídka zvnějšku). Rozlišují se potřeby a pudy, potřeby vrozené vs získané, primární vs sekundární (např. strach nebo touha po penězích), viscerogenní vs psychogenní, vnitřní vs vnější.
Zjednodušeně řečeno se v teoriích motivace jedná o vysvětlení lidského chování z hlediska vnitřních stavů a vnějších cílů (a jejich interakcí). Jak již bylo uvedeno, teorií motivace je mnoho [2,3]. Zatímco některé z nich kladou důraz na konfiguraci schopností, přání, potřeb, cílů a emocí, jiné (např. psychoanalýza) se zajímají o pudy a jejich konflikty, z nich vyplývající úzkost a obranné mechanizmy. Lze rozlišit tři obecné skupiny teorií motivace – hédonické, které se zabývají způsoby, jakými lidé hledají potěšení, čímž redukují svoji tenzi; kognitivní teorie, které k hédonickým motivům přidávají potřebu poznání; a růstové/aktualizační teorie. Teorie rozhodování s teoriemi motivace souvisejí dosti volně, je jich také několik skupin a v zásadě se jedná o snahu vysvětlit, proč se lidé přiklánějí k různým možnostem. Jsou značně komplexní a v tomto článku se věnujeme jen několika vybraným aspektům rozhodování.
Většina psychologických teorií motivace a rozhodování nepřináší vysvětlení souvisejících neurálních procesů a stejně tak neurovědní výzkumy se nevyjadřují k většině psychologických teorií motivace a rozhodování. Mezi oběma oblastmi výzkumu tak zatím neexistuje dostatečné propojení.
Zatímco fungování řeči, vnímání a paměti bylo studováno po mnoho desetiletí, možnosti studovat procesy motivace a rozhodování byly z důvodů jejich komplexnosti velmi omezeny. Přidružení těchto složitých duševních funkcí k prefrontální kůře bylo popsáno teprve v poslední době, a to díky nástupu nových technologií, především moderních zobrazovacích metod. Tyto technologie umožňují nahlédnout do činnosti mozku v souvislosti s chováním člověka. Metoda funkční magnetické rezonance (fMR) byla využita pro níže uvedené studie, které byly realizovány s cílem identifikovat ty struktury lidského mozku, jež by mohly být významně spojeny s procesy motivace. fMR umožňuje detekovat oblasti mozku, které se uplatňují při provádění konkrétní úlohy vyšetřovaným subjektem. Neurální aktivita je zde mapována pouze nepřímo, v návaznosti na lokální změnu oxygenace a perfuze mozkové kůry [4]. Existuje těsný vztah mezi lokální neuronální aktivitou a krevním průtokem danou oblastí. Synaptická aktivita zvyšuje krevní průtok. Při změnách množství kyslíku se mění poměr mezi oxygenovaným a deoxygenovaným hemoglobinem. Pokud se v konkrétním místě nahromadí větší množství oxyhemoglobinu, zaznamenáme zde silnější MR signál, než kdyby v této oblasti převažoval deoxyhemoglobin. Získaná data ukazují, které struktury byly během experimentu aktivovány, podílely se tak na realizaci zadaného úkolu a jsou asociovány s daným procesem, v našem případě s procesy motivace a rozhodování. Aktivní oblasti jsou vyznačeny na obr. 1.
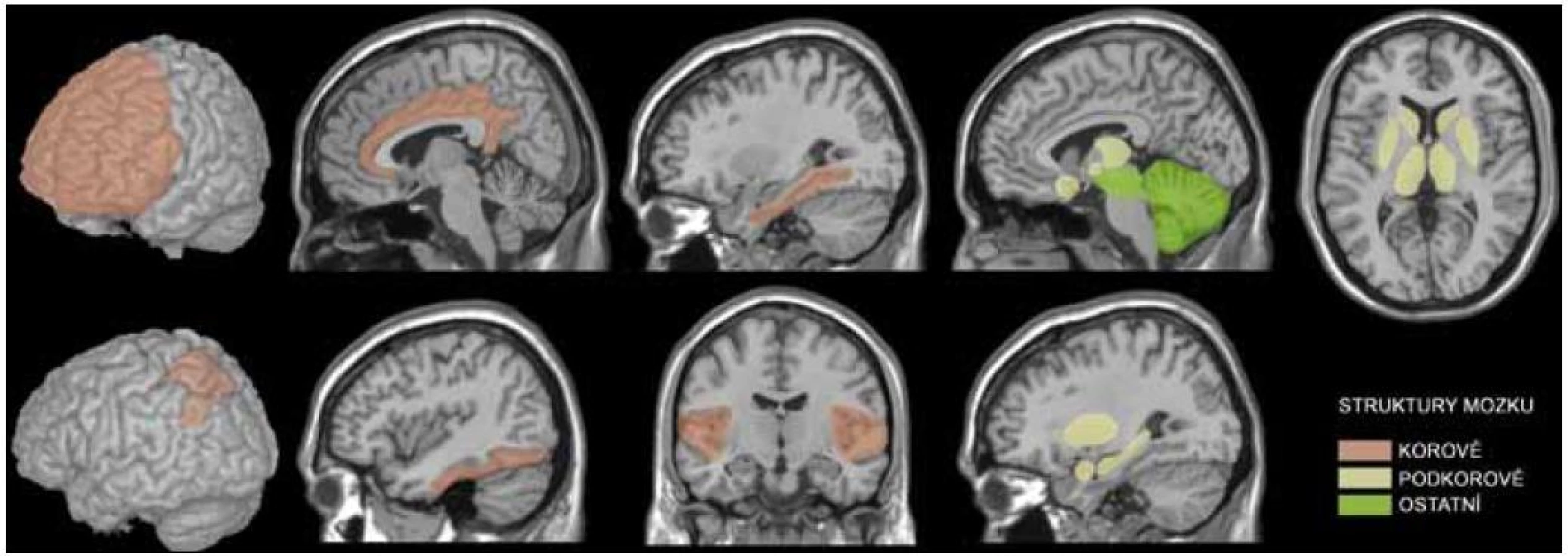
Motivace ziskem
Walter et al [5] zkoumali motivaci ziskem/odměnou (popř. ztrátou/trestem). Bylo zjištěno, že u člověka existuje systém podílející se na řízení chování v situacích, kdy lze očekávat zisk, který zahrnuje striatum, orbitofrontální kortex (OFC) a amygdalu. Nishimura et al [6] provedli recentně na skupině dobrovolníků experiment zaměřený na objasnění vztahu mezi neurálními mechanizmy a jednáním na základě instrukce, ale bez odměny. U zkoumaných subjektů byla při plnění úkolu bez odměny vyvolána aktivita v paralimbických oblastech (OFC, přední inzuly, laterálního prefrontálního kortexu, přední části cingula) a ve striatu. Zjištěné výsledky prokázaly, že tyto oblasti mozku plní významné role v motivovaném jednání. Také jsou zde vidět určité rozdíly mezi procesy motivace uplatňujícími se při motivaci ziskem oproti motivaci bez odměny.
Předmětem další práce [7] se stal výzkum neurálních odpovědí na odměnu nebo trest. Provádění úkolů zaměřených na získání odměny bylo v případě jejího dosažení spojeno s aktivací v mediálním OFC, v přední části gyrus cinguli, v amygdale a nucleus accumbens (NAcc). U trestů byla pozorována neurální odpověď v oblasti mediálního a dolního prefrontálního kortexu, horního parietálního kortexu a v inzule. Vnější motivace, při které je chování a prožívání stimulováno zvenčí, pozitivně korelovala se zvýšenými neurálními odpověďmi na odměnu v oblastech gyrus cinguli, amygdaly a putamen. Byl pozorován také negativní vztah mezi vnitřní motivací (např. potřebou, zájmem) a aktivací v těchto oblastech. Z výsledků této studie vyplývá, že neurální odpověď významně závisí na druhu motivace.
Zjištění z předchozí studie je podporováno výsledky Simona et al [8], kteří předpokládali, že variabilita v neurálním zpracování motivace pozorovaná u zdravých subjektů může odrážet individuální osobnostní rozdíly. Dobrovolníci s aktivnějším přístupem vykazovali během obdržení odměny více aktivity v oblasti ventrálního striata (VS). Také zde byla zjištěna větší aktivace v mediální orbitofrontální oblasti, která byla pozorována během obdržení i vynechání odměny. Subjekty s vyšší behaviorální inhibicí vykazovaly při obdržení odměny nižší aktivaci VS. Tyto nálezy naznačují, že tendence dosáhnout na odměnu nebo naopak vyhýbání se situacím spojeným s odměnou ukazují na zřetelnou souvislost s neurálním zpracováním těchto podnětů. U vyšetřovaných dobrovolníků s aktivnějším přístupem se projevovala vyšší senzitivita vůči pozitivním výsledkům a také menší tendence k vyhýbání se situacím s možností dosáhnout odměny, zatímco u dobrovolníků s vyšší behaviorální inhibicí byla zjištěna oslabená reakce na odměnu.
Další studie [9] zkoumala interakce mezi prefrontálními oblastmi lidského mozku (laterální OFC, mediální OFC a dorzolaterální prefrontální kortex, DLPFC), které doprovázejí vliv motivace na paměť. Dobrovolníci vykonali úlohu skládající se ze dvou úrovní. „Nízká motivace“ nebyla spojena s finanční odměnou, zatímco „vysoká motivace“ zahrnovala pravděpodobnost výhry jistého množství peněz. V části týkající se „nízké motivace“ ovlivnil OFC obou mozkových hemisfér pozitivně oblast levého DLPFC. Za podmínek „vysoké motivace“ se spojení v síti zahrnující oblast pravého OFC a oblast levého DLPFC měnilo od pozitivního k negativnímu, zatímco funkční konektivita v síti tvořené oblastí levého OFC a levého DLPFC byla ve srovnání s „nízkou motivací“ mírně zvýšena. Signifikantní na míře motivace závislou změnu vykázalo pouze funkční propojení mezi pravým laterálním OFC a levým DLPFC. Podle autorů se tato změna zdá být funkčním korelátem vlivu motivace na verbální pracovní paměť.
Peníze jsou často používaným prostředkem sloužícím jako odměna a zdroj motivace. S výzkumem motivačních mechanizmů souvisí i nová vědní disciplína neuroekonomie, spojující ekonomii, neurovědy a psychologii [10]. Jejím cílem je porozumět neurobiologickým mechanizmům, které se uplatňují při ekonomických volbách.
Knutson et al [11] ve své studii poukazují na aktivitu konkrétních struktur zapojených do rozhodovacích procesů během nakupování. Dobrovolníkům bylo nabízeno zboží různé cenové hodnoty. Při očekávání finančních zisků (výhodnosti nákupu) byla zjištěna aktivace v oblasti NAcc, zatímco v případech příliš vysoké ceny zboží došlo k aktivaci inzuly a současně ke snížení aktivity mesiálního prefrontálního kortexu (MPFC). Tyto poznatky naznačují, že aktivita zmíněných oblastí předchází a podporuje výsledek konečného rozhodnutí.
Kuhnen et al [12] zkoumali neurální aktivitu v mozku probíhající během rozhodování se o různých finančních investicích. Zjistili, že zatímco aktivace NAcc předcházela riskantnější volby, aktivace v oblasti inzuly souvisela s opatrnějšími volbami. Tyto výsledky se prakticky shodují s předchozí studií, která poukazuje na aktivaci NAcc v případě očekávání zisku, a tedy riskantnější volby, a aktivaci v oblasti inzuly při očekávání ztráty vedoucí k opatrnějším volbám. Aktivitou NAcc ve vztahu k očekávání odměn a trestů se zabývali i Knutson et al [13]. Jejich výsledky ukazují, že pokud zkoumaní dobrovolníci očekávali odměnu a udávali příjemný až blažený pocit, pak se zvýšila aktivita v oblasti NAcc. Pokud naopak předpokládali trest či finanční ztrátu, zkoumaná oblast zvýšenou aktivitu nevykazovala. Stejná skupina autorů [14] také v další studii poukázala na zvýšenou aktivitu MPFC, která byla zjištěna u dobrovolníků po získání finanční odměny, zatímco při jejím očekávání vykazovala zvýšení aktivity oblast VS. Pokud dobrovolníci získali očekávanou finanční odměnu, aktivita v oblasti MPFC vykazovala zvýšení, zatímco pokud očekávanou odměnu neobdrželi, aktivita zde poklesla.
Morální motivace
Další zajímavou oblastí je studium motivačních procesů, které souvisejí s chováním podle určitých morálních principů. Klinické a experimentální studie začaly poskytovat důkazy o vlivu kulturních a biologických faktorů na lidskou morálku. Nedávné výzkumy naznačují, že vysoce kulturně závislé osobnostní rysy projevující se při (ne)morálním jednání, závisejí na integritě prefrontální mozkové kůry [15]. U násilných zločinců a lidí postrádajících schopnost hodnocení na základě etických hledisek (schopnost říci, co je v hypotetické situaci správné, a co ne) byl popsán neobvykle vysoký výskyt poškození nebo dysfunkcí čelního laloku. Kromě jiného umožnila fMR zkoumat cerebrální procesy, ke kterým dochází v průběhu morálního rozhodování.
Tak vznikla například i studie, při níž dobrovolníci četli sérii krátkých příběhů obsahujících morální aspekty, které u nich vyvolaly různé emoce včetně odporu a rozhořčení. Negativní emoce byly spojeny s aktivací regionů frontálních a temporálních laloků. Jednalo se zejména o oblast OFC, k jejíž aktivaci došlo bilaterálně. V levé hemisféře byla aktivace rozšířena i na gyrus frontalis inferior. K aktivaci došlo také v oblastech levého frontálního opercula a bilaterálně u gyrus frontalis superior. Tato práce vyzdvihuje důležitost prefrontální a orbitofrontální mozkové kůry při tvorbě morálního úsudku [16]. V následující studii byly u dobrovolníků vyvolány specifické emoce, aby bylo možné sledovat, které oblasti mozku budou aktivovány. Za tímto účelem byly použity konkrétní scénáře pro vyvolání různých emocí, jako jsou vina, rozhořčení a soucit [17]. Různé kategorie emocí vedly k aktivaci rozdílných oblastí. Prožívání viny, rozpaků a soucitu souviselo s aktivací oblasti předního mediálního prefrontálního kortexu a sulcus temporalis superior, při zážitcích empatie došlo k dodatečnému zapojení mezolimbické dráhy. Hodnotící emoce, jako jsou odpor nebo rozhořčení, byly asociovány s aktivací amygdaly, parahipokampální a fuziformní oblasti.
Studii, ve které si museli dobrovolníci vybírat z nabídnutých scénářů, a byli tak postaveni před morální dilema, uskutečnili Schaich Borg et al [18]. Účastníkům studie byly předloženy scénáře zahrnující volbu mezi následky s různou mírou škody. Autoři rozlišovali tzv. morální scénáře, které obsahovaly volby mezi různou mírou negativních důsledků pro lidské účastníky, a tzv. ne-morální scénáře, které se týkaly negativních důsledků pro věci nebo majetek. Morální scénáře vyvolaly aktivitu v podobných oblastech mozku jako obdobné ne-morální scénáře. Ve srovnání s ne-morálními scénáři vyvolaly morální scénáře více aktivity v oblastech asociovaných s kognitivními funkcemi (jako je DLPFC) a méně aktivity v oblastech asociovaných s emocemi (OFC, temporální lalok). Srovnáno s obdobnými ne-morálními scénáři, dilema umožňující volbu minimalizující škody vyvolalo více aktivity v oblastech asociovaných s emocemi (OFC a temporální lalok) a méně aktivity v oblastech asociovaných s kognicí (zahrnující gyrus angularis a gyrus frontalis superior). A konečně ve srovnání se scénáři zahrnujícími pouze neúmyslné poškození vyvolávají scénáře způsobující úmyslné poškození více aktivity v oblastech asociovaných s emocemi (OFC a temporální lalok) a méně aktivity v oblastech asociovaných s kognicí (zahrnující gyrus angularis a gyrus frontalis superior). Z těchto výsledků lze vyvozovat, že různé morální úsudky jsou podporovány rozlišitelnými neurálními systémy.
Otázka, co motivuje jedince chovat se morálně, je předmětem mnoha dalších vědních disciplín, například filozofie, psychologie nebo i trestního práva. Tyto motivy jsou hluboce ovlivněny sociálním učením a individuálními biologickými rozdíly [19]. Odpovědi mohou přispět k pochopení, jak může lidská mysl produkovat antisociální chování, jako je zpronevěra, podvody, krádeže, přepadení, znásilnění a vraždy a pomoci posoudit otázky trestní zodpovědnosti za bezprostřední problémy řešené v dnešní době v soudních síních [20].
Rozhodovací procesy
Jak výběr z alternativ vedoucích k zisku/odměně nebo ztrátě/trestu, tak řešení morálních dilemat předpokládá volbu ze dvou a více možností, tzn. nějakou formu rozhodování. Proces výběru odpovědi z několika alternativ byl zkoumán ve studii Pauluse a Franka [21]. Během výběru konkrétní alternativy byla zaznamenána výrazně vyšší aktivace v gyrus frontalis medialis, k aktivaci došlo i v oblastech posteriorního parietálního kortexu, předního cingula a levé přední inzuly. V jiné studii bylo také srovnáváno emoční a kognitivní hledisko volby [22]. Při výběru z emočního hlediska byla aktivována přední inzula, operculum a přední cingulum, zatímco při rozhodnutích kognitivního hlediska byl více aktivní DLPFC. Aktivace v oblasti předního cingula korelovala se subjektivním oceněním stimulu. Oblasti mediálního OFC a ventrolaterálního prefrontálního kortexu (PFC) byly aktivnější, pokud emoční stimuly vedly k výběru. Na základě výsledků této studie se soudí, že subjektivní preference rozhodnutí je ovlivňována oblastmi inzuly a cingulátního kortexu, zatímco oblasti mediálního OFC, ventrolaterálního PFC a inzuly přispívají k vyhodnocení podnětů a motivačních aspektů výběru odpovědi.
Vzhledem k tomu, že žijeme ve složitých sociálních podmínkách, je značná část našich rozhodnutí ovlivňována sociálními interakcemi. Významnou úlohu zde hraje zejména PFC, jehož funkce souvisí se zohledňováním sociálních interakcí během rozhodování [23]. V souvislosti s lézemi PFC jsou často popisovány behaviorální poruchy a poruchy sociální percepce. Tuto skutečnost podporují i výsledky studie porovnávající skupinu pacientů s poškozením PFC se zdravými dobrovolníky [24]. Obě skupiny sledovaly nahrané sociální interakce založené na neverbálních nápovědách a odhadovaly interpersonální vztahy osob zúčastněných v nahrané scénce, jako např. stupeň intimity mezi dvěma osobami. Pacienti s lézí OFC vykazovali sníženou sociální percepci oproti zdravým dobrovolníkům. Obdobné výsledky byly stejným způsobem získány i u pacientů s poškozením DLPFC. Tato zjištění potvrzují význam PFC v procesech uplatňovaných během sociální percepce.
Poruchy motivace
Poruchy motivace mohou souviset s více mozkovými strukturami. Všechny tyto poruchy souvisí s poškozením nebo dysfunkcí prefrontální mozkové kůry a dalších struktur, zvláště těch, které jsou s prefrontální kůrou úzce spojeny (např. přední cingulum, neostriatum, dorzomediální thalamické jádro, ventrální tegmentální area (VTA), mezencefalon), nebo jejich vzájemných drah [15]. Během běžného vyšetření bývá často obtížné změny motivačních procesů postřehnout.
V klinické praxi je patrně nejlépe rozpoznatelnou změnou apatie. Apatie se vyskytuje často jako příznak dalších syndromů, ale i jako syndrom sám o sobě [25]. Je popisována jako ztráta motivace a zhoršení behaviorálních, kognitivních a emočních aspektů při nezměněném vědomí [1]. Typicky je apatie spojena se strukturním postižením mesiálních prefrontálních oblastí. Apatie je však složitějším klinickým příznakem, který může být také projevem mnoha psychiatrických onemocnění a psychosociálních patologií [26].
Pokles motivace také patří mezi první příznaky, které se objevují u většiny demencí, včetně demence Alzheimerova typu [15]. Cílem recentní studie Dujardinové et al [27] bylo prověřit u pacientů s demencí u Parkinsonovy nemoci, zda se pokles kognitivních funkcí objevil častěji u apatických jedinců než u subjektů, které apatii nevykazovaly. Při vstupu do studie měli pacienti vykazující apatii oproti neapatickým subjektům výrazně nižší globální kognitivní status. Po 18 měsících bylo zjištěno, že apatická skupina vykazovala výrazně vyšší rychlost změn příznačných pro demenci než skupina apatii nejevící. Dokonce i v případě pacientů, kteří demenci nevykazovali, docházelo k poklesu kognitivní výkonnosti mnohem častěji u apatických pacientů než u těch, v jejichž případech nebyla apatie pozorována. Tyto poznatky naznačují, že u pacientů trpících Parkinsonovou nemocí nevykazujících demenci nebo depresi může být apatie prediktivním faktorem pro pokles kognitivních funkcí a rozvoj demence.
Běžné motivační mechanizmy nefungují zpravidla ani u autistických osob. Odlišnosti ve struktuře a funkci CNS autistických jedinců jsou patrné jak na mikroskopické, tak makroskopické úrovni. Přestože popsané nálezy nejsou konzistentní, v centru pozornosti stojí frontální, temporální a parietální kortex, bazální ganglia (zejména nucleus caudatus), amygdala, hipokampus, thalamus a mozeček. Makroskopické abnormality v těchto oblastech jsou jak ve smyslu zvýšení, tak i snížení objemu popsaných struktur [28]. Kleinhans et al [29] vyšetřili dobrovolníky s autizmem MR spektroskopií a zjistili významné snížení NAA (N-acetylaspartát) ve všech oblastech mozku. V souladu s tím výzkumy uskutečněné s pomocí protonové magnetické rezonanční spektroskopie (1H MRS) potvrdily, že pacienti postižení cévní mozkovou příhodou (CMP) a trpící následně apatií měli nižší podíl NAA/Cr (N-acetylaspartát//kreatin) v pravém frontálním laloku oproti neapatickým subjektům [30]. Dále autoři srovnávali dvě skupiny pacientů podle toho, byla-li postižena ischemií levá, nebo pravá hemisféra. U skupiny apatických pacientů s levostranným poškozením byl nalezen signifikantně nižší poměr NAA/Cr v oblasti pravé mozkové hemisféry. Tyto výsledky směřují k závěru, že v některých případech může apatie vyskytující se po CMP souviset s metabolickými změnami v oblastech mozku vztahujícími se k motivačním procesům, a to i když tyto oblasti nebyly přímo zasaženy ischemií.
Oslabení motivace bývá pozorováno i u pacientů s traumatickým poraněním mozku [1]. Poškození struktur popsaných v předchozí části článku, tzn. prefrontální, oblasti předního cingula, NAcc, VS, thalamu a VTA, může způsobovat poruchy motivace. Typickými příznaky jsou pak apatie, abulie a totální absence spontánního chování. Lidské chování se stává bezúčelným, chaotickým a chudým, a to i když postiženému zůstane zachována většina kognitivních schopností. K něčemu podobnému zřejmě došlo i u literárně slavného pacienta P. P. Gage v polovině 19. století. Jde o dobře zdokumentovaný případ těžkého otevřeného poranění mozku masivní ocelovou tyčí s poškozením obou mesiálních prefrontálních oblastí. Gage se dokázal překvapivě rychle z extrémního zranění zotavit. Jeho hybnost i většina kognitivních funkcí zůstaly neporušeny. Došlo u něj ale k výrazným osobnostním změnám. Odpovědný, inteligentní, společensky dobře přizpůsobivý, výkonný a svědomitý muž se po incidentu zásadním způsobem změnil. Stal se nerozhodným a neschopným držet se svých plánů [31]. Přestal dodržovat svoje závazky a v důsledku toho opakovaně přišel i o práci. Zasažené oblasti tedy podle všeho hrály důležitou roli v jeho procesech motivace a rozhodování.
Další studie zkoumala podobnosti a rozdíly u apatií vyvolaných různými patologickými procesy. Byly srovnávány následné stavy po traumatickém poranění mozku s obrazem apatie u pacientů trpících schizofrenií [32]. U obou skupin byla zjištěna stejná míra apatie, ale pacienti trpící schizofrenií vykazovali vážnější příznaky anhedonie, alogie a menší schopnosti prožitků. Juckel et al [33] použili fMR, aby sledovali odpověď ve VS u nemedikovaných schizofreniků při prezentaci podnětů naznačujících budoucí zisk a ztrátu. Výsledky odhalily sníženou aktivaci v levém VS při prezentaci odměny v porovnání se zdravými jedinci. Tato reakce ve VS při porovnání odměna vs neutrální vizuální podnět nepřímo korelovala se závažností negativních příznaků. V těchto případech může vysoká hladina dopaminu v oblasti VS narušit neurální zpracování podnětů týkajících se predikce odměny. A to pak může přispět k negativním symptomům, jako jsou anhedonie, apatie a ztráta motivace. Další klinické studie naznačují, že u medikovaných schizofreniků jsou některá netypicky používaná neuroleptika (např. olanzapin, risperidon) efektivnější než typická neuroleptika (např. haloperidol) v redukci negativních symptomů, které zahrnují apatii a anhedonii [34]. Schizofrenici léčení atypickými neuroleptiky vykazovali v odpovědi na podněty indikující odměnu aktivaci pravého VS, zatímco skupina schizofreniků léčená typickými neuroleptiky signifikantně zvýšenou odpověď nevykazovala. U zdravých dobrovolníků byla zjištěna aktivace VS bilaterálně. U pacientů léčených typickými neuroleptiky korelovalo snížení aktivace v levém VS se závažností negativních symptomů.
V posledních letech jsou poměrně často popisovány izolované poruchy motivace a jednání u neurologických pacientů, které mohou být podle všeho způsobeny bilaterální lézí v bazálních gangliích [35]. Tyto léze mohou být ischemické nebo toxické. Usuzuje se, že dochází k bilaterální dysfunkci kortiko-subkortikálních okruhů, která se klinicky projevuje dramatickým snížením spontánního jednání postižených jedinců. Dochází k poklesu nebo absenci spontánních myšlenek. V důsledku toho může ztráta zájmu u těchto pacientů ovlivnit jejich spolupráci při diagnostice. Habib [36] publikoval pozorování dvou jinak zdravých jedinců, u nichž se rozvinuly výrazné poruchy motivace. U popsaných případů docházelo mimo jiné k výraznému poklesu vůle, aktivity, schopnosti spontánně myslet a k hluboké netečnosti. S pomocí SPECT byla v prvním případě zjištěna bilaterální hypoperfuze v oblasti bazálních ganglií bez významných změn kortikálního krevního průtoku a v dalším případě pak bylo prokázáno bilaterální poškození hlavy nucleus caudatus. Oba pacienti měli společné nejen dramatické změny chování, ale také velmi podobně lokalizované léze. Lakunární infarkty byly u obou pacientů nejvíce vyjádřeny v oblasti nucleus caudatus. Popsaný syndrom je v současnosti popisován jako athymhormia.
Testována byla také hypotéza, že apatie vyvolaná hlubokou mozkovou stimulací subtalamického jádra u Parkinsonovy nemoci koreluje se změnami v metabolizmu glukózy [37]. Dvanáct pacientů s Parkinsonovou nemocí bylo s pomocí 18FDG-PET hodnoceno tři měsíce před stimulací a tři měsíce po ní. Po třech měsících od stimulace došlo k výraznému prohloubení apatie. Mezi apatií a změnou metabolizmu glukózy byla pozorována pozitivní korelace, a to zejména v pravém gyru frontalis medialis (BA 10) a v pravém gyru frontalis inferior (BA 46, BA 47). Negativní korelace byla pozorována v pravém zadním cingulu (BA 31) a levém mediálním čelním laloku (BA 9). Tyto předběžné výsledky naznačují, že subthalamické jádro je jednou z klíčových struktur ovlivňujících motivační mechanizmy.
Anhedonie a pokles motivace patří mezi symptomy deprese. Neurobiologické výzkumy naznačují, že u tohoto v současnosti poměrně častého onemocnění může hrát roli narušení frontálních dopaminergních projekcí. Deprese často limituje efektivní léčbu motorických příznaků, snižuje funkční schopnosti nemocných a zhoršuje kvalitu jejich života [38]. Popisovány jsou také souvislosti mezi depresí a nadměrným užíváním alkoholu nebo drog.
Obecně se má za to, že alkohol ovlivňuje emoce, jako jsou úzkost a strach, pomocí modulace zpracování informací. To potvrzuje také studie zkoumající akutní účinky alkoholu na afektivní zpracování obrazových stimulů pomocí metody evokovaných potenciálů (ERP) [39]. Dvě skupiny dobrovolníků (první skupina účastníků obdržela nápoj obsahující mírné dávky alkoholu, zatímco druhé skupině byl podán nealkoholický nápoj) dostaly za úkol pozorovat obrazové podněty s příjemnou, nepříjemnou či neutrální náplní. Výsledky ukazují, že alkohol snižuje mozkovou činnost při sledování nepříjemné informace a potvrzují vliv alkoholu na zpracování afektivních informací. Existuje několik rozličných metabolických cest vzniku závislosti na alkoholu. Každá z nich je zprostředkována zvláštními nervovými spoji a drahami. Bylo prokázáno, že alkohol působí na tvorbu endorfinů a výrazně zvyšuje hladinu dopaminu v některých oblastech mozku (NAcc) a v mozkových drahách, způsobem podobným působení některých drog.
Již poměrně dlouhou dobu se v psychiatrii diskutuje o tzv. amotivačním syndromu, snad souvisejícím s negativním vlivem užívání marihuany na motivaci. Tento syndrom se projevuje apatií, ztrátou energie, snížením pozornosti a zhoršením pracovních výkonů. Nestor et al [40] recentně uskutečnili výzkum prověřující chronické účinky marihuany na ty oblasti mozku, které zpracovávají odměnu a mají vliv na motivaci člověka. U uživatelů konopí byla s pomocí fMR zjištěna signifikantně zvýšená odpověď v oblasti pravého VS během očekávání zisku, která signifikantně korelovala s délkou a celkovým množstvím užité drogy. V levém inzulárním kortexu naopak došlo ke snížení aktivity jako reakce na ztrátu a zkoumané subjekty v souladu s tímto nálezem vykazovaly sníženou snahu vyhnout se ztrátě. Také Martin-Soelch et al [41] ve své studii potvrzují, že pravidelné užívání konopí se dotýká některých aspektů motivace a že jak nikotinizmus, tak užívání konopí vede k podobným změnám v motivačních procesech. Zde autoři srovnávali motivaci u uživatelů marihuany, pravidelných kuřáků tabáku a nekuřáků během plnění úkolu, při němž byli dobrovolníci motivováni finančním ziskem. U skupiny nekuřáků byla zjištěna významná pozitivní korelace mezi náladou subjektů a ziskem peněžní odměny, ale zbylé dvě skupiny tuto závislost nevykazovaly. Užívání konopí však ovlivnilo motivaci mnohem výrazněji než kouření tabáku. Předpokládá se, že jsou za to odpovědné změny v hladině dopaminu a poruchy v neurálních okruzích podílejících se na motivaci a pozornosti.
Velmi zajímavý je výzkum tzv. extrémní lenosti, která může podle australských vědců souviset se zdravotními důvody. Motivační deficit, charakterizovaný ohromující a vysilující apatií, byl podle citované studie objeven u každého pátého vyšetřovaného jedince ze skupiny testovaných Australanů [42]. V extrémních případech může být tato porucha pro postiženého dokonce fatální. Onemocnění lze diagnostikovat pomocí testů hodnotících stupeň motivace v kombinaci s pozitronovou emisní tomografií. V současnosti je tato patologie nedostatečně prozkoumána a diagnostikována.
Možnosti terapie
Léčba poruch motivačních procesů závisí na příčině, se kterou tyto defekty souvisí. Přesná diagnóza se stává zvláště důležitou s příchodem stále více cílených terapií, které mohu být farmakologické, kognitivně-rehabilitační a další. Na zmírnění apatie jsou podávány léky zvyšující hladinu dopaminu (amantadin, amfetamin, bromocriptin, bupropion, metylfenidát a selegilin) [25,38]. Také je možné použít preparáty stimulující CNS (pemolin) [38]. Obecně lze doporučit využití poradenství s podporou systému vnitřní motivace a spojovat podněty s odměnou. U většiny vážnějších případů však bohužel dosavadní možnosti léčby nezaznamenaly trvalejší úspěch.
Struktury uplatňující se v motivačních procesech
Snahou současného široce koncipovaného neurovědního výzkumu je mimo jiné lépe porozumět souvislostem mezi cerebrálními komponentami a konkrétními mentálními procesy. Tento souhrnný článek zaměřený na studium lidského mozku v souvislosti s motivačními a rozhodovacími procesy uvádí řadu recentních prací, jejichž výsledky identifikují konkrétní cerebrální struktury ve vztahu k motivačním mechanizmům (NAcc, inzula, OFC, DLPFC, VS, přední cingulum, amygdala). Pokusili jsme se přehledně je uspořádat v tab. 1. Obsahuje jednotlivé struktury mozku členěné pokud možno hierarchicky spolu se stručným odkazem na výzkumné výsledky a citované studie. Byla také zjištěna souvislost mezi hladinou dopaminu, integritou dopaminergního systému a strukturami mozku zpracovávajícími odměny a ovlivňujícími motivační procesy [25,33,40,43,44]. Významnou roli může v některých případech zřejmě hrát i snížená hladina NAA [29,30], která však na druhé straně může být pouhým epifenoménem, odrážejícím pouze v obecné rovině dysfunkci dané oblasti.
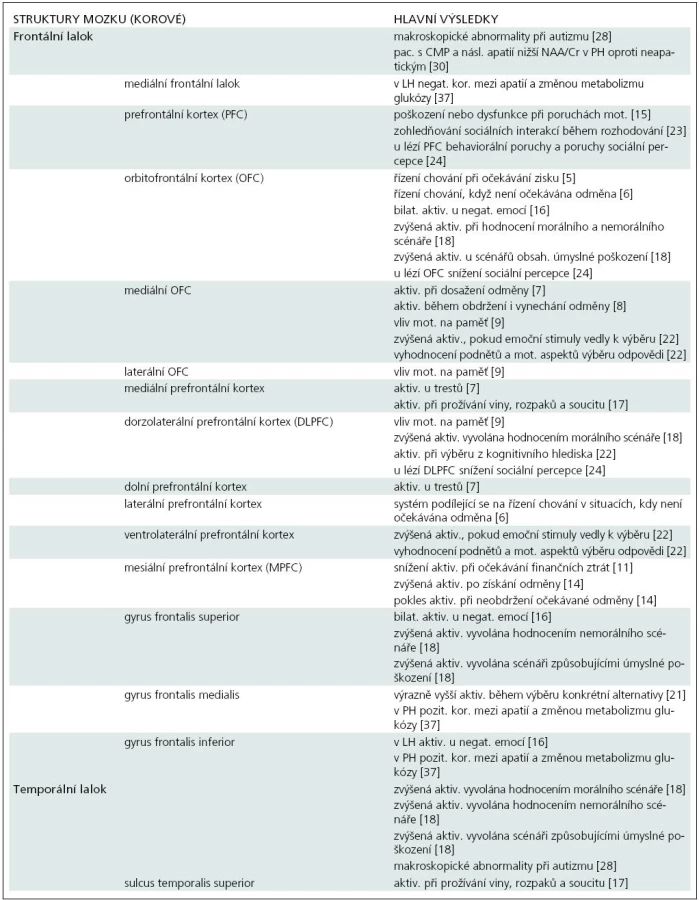


Léze uvedených cerebrálních struktur pak mohou souviset s poruchami motivace u postižených jedinců. Deficit motivace může být obecně způsoben širokou škálou neurologických, psychiatrických a geriatrických poruch. Přesná diagnóza se stává zvláště důležitou s příchodem stále více cílených terapií. Poruchy motivace mají velmi negativní dopad na kvalitu života pacienta, jeho produktivitu, možnost vést normální život a na jeho úlohu ve společnosti.
Práce byla uskutečněna za podpory výzkumných záměrů MŠMT ČR č. MSM0021622404 (MB) a AV0Z70250504 (TU).
Mgr.
Jana Zelinková
I.
neurologická klinika
LF
MU a FN u sv. Anny
Pekařská
53
656
91 Brno
e-mail:
85355@mail.muni.cz
Přijato
k recenzi: 16. 8. 2010
Přijato
do tisku: 7. 12. 2010
Zdroje
1. Marin RS, Wilkosz PA. Disorders of diminished motivation. J Head Trauma Rehab 2005; 20(4): 377–388.
2. Madsen KB. Moderní teorie motivace. Praha: Academia 1979.
3. Madsen KB. Teorie motivace. Praha: Academia 1972.
4. Chlebus P, Mikl M, Brázdil M, Krupa P. Funkční magnetická rezonance – úvod do problematiky. Neurol pro praxi 2005; 6(3): 133–139.
5. Walter H, Abler B, Ciaramidaro A, Erk S. Motivating forces of human actions: Neuroimaging reward and social interaction. Brain Res Bull 2005; 67(5): 368–381.
6. Nishimura M, Yoshii Y, Watanabe J, Ishiuchi S. Paralimbic system and striatum are involved in motivational behavior. Neuroreport 2009; 20(16): 1407–1413.
7. Linke J, Kirsch P, King AV, Gass A, Hennerici MG, Bongers A et al. Motivational orientation modulates the neural response to reward. Neuroimage 2010; 49(3): 2618–2625.
8. Simon JJ, Walther S, Fiebach CJ, Friederich HC, Stippich C, Weisbrod M et al. Neural reward processing is modulated by approach- and avoidance-related personality traits. Neuroimage 2010; 49(2): 1868–1874.
9. Szatkowska I, Bogorodzki P, Wolak T, Marchewka A, Szeszkowski W. The effect of motivation on working memory: An fMRI and SEM study. Neurobiol Learn Mem 2008; 90(2): 475–478.
10. Glimcher PW, Rustichini A. Neuroeconomics: the consilience of brain and decision. Science 2004; 306(5695): 447–452.
11. Knutson B, Rick S, Wimmer GE, Prelec D, Loewenstein G. Neural predictors of purchases. Neuron 2007; 53(1): 147–156.
12. Kuhnen CM, Knutson B. The neural basis of financial risk taking. Neuron 2005; 47(5): 763–770.
13. Knutson B, Adams CM, Fong GW, Hommer D. Anticipation of increasing monetary reward selectively recruits nucleus accumbens. J Neurosci 2001; 21(16): RC159.
14. Knutson B, Fong GW, Bennett SM, Adams CM, Hommer D. A region of mesial prefrontal cortex tracks monetarily rewarding outcomes: characterization with rapid event-related fMRI. Neuroimage 2003; 18(2): 263–272.
15. Goldberg E. Attention and Motivation. In: Bloom FE, Beal MF, Kupfer DJ (eds). The Dana Guide to Brain Health. New York: Dana Press 2003: 196–199.
16. Moll J, de Oliveira-Souza R, Moll FT, Ignácio FA, Bramati IE, Caparelli-Dáquer EM et al. The moral affiliations of disgust. A functional MRI study. Cogn Behav Neurol 2005; 18(1): 68–78.
17. Moll J, de Oliveira-Souza R, Garrido GJ, Bramati IE, Caparelli-Daquer EM, Paiva ML et al. The self as a moral agent: Linking the neural bases of social agency and moral sensitivity. Soc Neurosci 2007; 2(3–4): 336–352.
18. Schaich Borg J, Hynes C, Horn JV, Grafton S, Sinnott-Armstrong W. Consequences, action and intention as factors in moral judgments: an fMRI investigation. J Cog Neurosci 2006; 18(5): 803–817.
19. Moll J, de Oliveira-Souza R, Zahn R. The neural basis of moral cognition: sentiments, concepts, and values. Ann NY Acad Sci 2008; 1124(1): 161–180.
20. Gazzaniga MS. The law and neuroscience. Neuron 2008; 60(3): 412–415.
21. Paulus MP, Frank LR. Ventromedial prefrontal cortex activation is critical for preference judgments. Neuroreport 2003; 14(10): 1311–1315.
22. Chaudhry AM, Parkinson JA, Hinton EC, Owen AM, Roberts AC. Preference judgements involve a network of structures within frontal, cingulate and insula cortices. Eur J Neurosci 2009; 29(5): 1047–1055.
23. Rilling JK, Sanfey AG. The neuroscience of social decision-making. Annu Rev Psychol 2011; 62: 23–48.
24. Mah L, Arnold MC, Grafman J. Impairment of social perception associated with lesions of the prefrontal cortex. Am J Psychiatry 2004; 161(7): 1247–1255.
25. Marin RS, Fogel BS, Hawkins J, Duffy J, Krupp B. Apathy: a treatable syndrome. J Neuropsychiatry Clin Neurosci 1995; 7(1): 23–30.
26. Marin RS, Chakravorty S. 18. Disorders of Diminished Motivation. In: Silver JM, McAllister TW, Yudofsky SC (eds). Textbook of Traumatic Brain Injury. Washington, DC: American Psychiatric Publishing 2005: 337–352.
27. Dujardin K, Sockeel P, Delliaux M, Destée A, Defebvre L. Apathy may herald cognitive decline and dementia in Parkinson’s disease. Mov Disorders 2009; 24(16): 2391–2397.
28. Fialová L. Dětský autizmus v pohledu genetiky a vývojové neurobiologie. Psychiatrie 2007; 11(4): 220–225.
29. Kleinhans NM, Schweinsburg BC, Cohen DN, Müller RA, Courchesne E. N-acetyl aspartate in autism spectrum disorders: regional effects and relationship to fMRI activation. Brain Res 2007; 1162: 85–97.
30. Glodzik-Sobanska L, Slowik A, Kieltyka A, Kozub J, Sobiecka B, Urbanik A et al. Reduced prefrontal N--acetylaspartate in stroke patients with apathy. J Neurol Sci 2005; 238(1–2): 19–24.
31. Damasio H, Grabowski T, Frank R, Galaburda AM, Damasio AR. The return of Phineas gage: clues about the brain from the skull of a famous patient. Science 1994; 264(5162): 1102–1105.
32. Rao V, Spiro JR, Schretlen DJ, Cascella NG. Apathy Syndrome After Traumatic Brain Injury Compared With Deficits In Schizophrenia. Psychosomatics 2007; 48(3): 217–222.
33. Juckel G, Schlagenhauf F, Koslowski M, Wüstenberg T, Villringer A, Knutson B et al. Dysfunction of ventral striatal reward prediction in schizophrenia. Neuroimage 2006; 29(2): 409–416.
34. Juckel G, Schlagenhauf F, Koslowski M, Filonov D, Wüstenberg T, Villringer A et al. Dysfunction of ventral striatal reward prediction in schizophrenic patients treated with typical, not atypical, neuroleptics. Psychopharmacology 2006; 187(2): 222–228.
35. Habib M. Activity and motivational disorders in neurology: proposal for an evaluation scale. Encephale 1995; 21(5): 563–570.
36. Habib M. Athymhormia and disorders of motivation in basal ganglia disease. J Neuropsychiatry Clin Neurosci 2004; 16(4): 509–524.
37. Le Jeune F, Drapier D, Bourguignon A, Péron J, Mesbah H, Drapier S et al. Subthalamic nucleus stimulation in Parkinson disease induces apathy: a PET study. Neurology 2009; 73(21): 1746–1751.
38. Anders M, Roth J, Uhrová T. Organická deprese při vybraných onemocněních mozku. Psychiatrie 2005; 9(1): 32–39.
39. Franken IH, Nijs IM, Muris P, Van Strien JW. Alcohol selectively reduces brain activity during the affective processing of negative information. Alcohol Clin Exp Res 2007; 31(6): 919–927.
40. Nestor L, Hester R, Garavan H. Increased ventral striatal BOLD activity during non-drug reward anticipation in cannabis users. Neuroimage 2010; 49(1): 1133–1143.
41. Martin-Soelch Ch, Kobel M, Stoecklin M, Michael T, Weber S, Krebs B et al. Reduced Response to Reward in Smokers and Cannabis Users. Neuropsychobiology 2009; 60(2): 94–103.
42. Moynihan R. Scientists find new disease: motivational deficiency disorder. BMJ 2006; 332: 745.
43. Menon V, Levitin DJ. The rewards of music listening: response and physiological connectivity of the mesolimbic system. Neuroimage 2005; 28(1): 175–184.
44. Volkow ND, Wang GJ, Kollins SH, Wigal TL, Newcorn JH, Telang F et al. Evaluating dopamine reward pathway in ADHD: clinical implications. JAMA 2009; 302(10): 1084–1091.
Štítky
Dětská neurologie Neurochirurgie NeurologieČlánek vyšel v časopise
Česká a slovenská neurologie a neurochirurgie
2011 Číslo 4
Nejčtenější v tomto čísle
- Porucha pozornosti s hyperaktivitou (attention deficit/hyperactivity disorder – ADHD)
- Opožděný akutní subdurální hematom
- Neurologické komplikace při onemocnění herpes zoster – kazuistika
- Vrozená myotonie na podkladě mutací v genu pro chloridový kanál ClC-1